
|
Malesian Euphorbiaceae Descriptions |
|
Stuppy, W., P.C. van Welzen, P. Klinratana & M.C.T. Posa. 1999. Revision of the genera Aleurites, Reutealis and Vernicia (Euphorbiaceae). Blumea 44: 73–98.
Goto on this page:
Aleurites J.R.Forst. & G.Forst., Char. Gen. Pl. (1776) 56; Müll.Arg. in DC., Prodr. 15, 2 (1866) 722; Hook.f., Fl. Brit. India 5 (1887) 384; Pax in Engl., Pflanzenr. IV.147 (1910) 128; J.J.Sm., Meded. Dept. Landb. Ned.-Indië 10 (1910) 550; Backer & Bakh.f., Fl. Java 1 (1963) 477; Airy Shaw, Kew. Bull. 20 (1967) 393; Kew Bull. 26 (1972) 213; Kew Bull. Add. Ser. 4 (1975) 29; E.Walker, Fl. Okinawa S. Ryukyo Isl. (1976) 644; Airy Shaw, Kew Bull. Add. Ser. 8 (1980) 25; Muelleria 4 (1980) 230; Kew Bull. 36 (1981) 252; Alph. Enum. Euphorb. Philipp. Isl. (1983): 4; W.L.Wagner, D.R.Herbst & Sohmer, Man. Fl. Pl. Hawai’i 1 (1990) 597; G.L.Webster, Ann. Missouri Bot. Gard. 81 (1994) 114; P.I.Forst., Muelleria 9 (1996) 6; Stuppy et al., Blumea 44: 79; Radcl.-Sm., Gen. Euphorbiacearum (2001) 330; Stuppy et al. in Chayam. & Welzen, Fl. Thailand 8, 1 (2005) 49; G.L.Webster in Kubitzki, Fam. Gen. Vasc. Pl. 11 (2014) 183. — Aleurites J.R.Forst. & G.Forst. section Camirium (Rumph. ex Gaertn.) Müll.Arg. in DC., Prodr. 15, 2 (1966) 723, nom. inval.; Pax in Engl., Pflanzenr. IV.147 (1910) 129. — Type: Aleurites triloba J.R.Forst. & G.Forst. [= Aleurites moluccana (L.) Willd.]
Camirium Rumph. ex Gaertn., Fruct. Sem. Pl. 2 (1791) 194. — Type: Camirium cordifolium Gaertn. [= Aleurites moluccana (L.) Willd.]
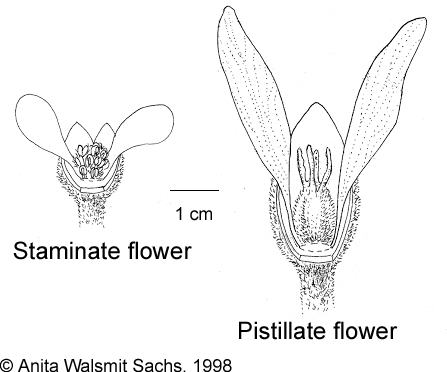
Trees, monoecious, evergreen. Indumentum of greyish or fulvous-silver to ferruginous-silver stellate, multicellular hairs, glandular or stinging hairs absent. Young twigs terete, scurfily stellate pubescent. Stipules minute, cylindric, stellate-pubescent, early caducous, leaving two small circular scars. Petiole terete, slightly ribbed or striate, evenly scurfily or shortly stellate-pubescent or puberulous, with two (rarely three) sessile, discoid, shining glands adaxially at junction with blade, the glands exudating a sweet sap. Leaves alternate, simple, ovate to ovate-lanceolate or ovate-trullate, not or shallowly 3- or 5-palmately lobed, without glands in marginal sinuses between lobes, densely stellate-pubescent above and beneath when young, glabrescent; lobes triangular-deltoid; base cuneate, rounded, truncate, or (especially if the leaves are lobed) shallowly to moderately cordate or reniform; margin entire; apex and lobes acute to acuminate; basally palmately 3- or 5-veined, pinnate along the midrib with 4–8 major lateral veins per side. Inflorescences terminal, solitary, many-flowered pyramidal thyrses, conical, branching from the base, bisexual or rarely unisexual with the flowers in bracteate clusters, protogynous with a solitary pistillate flower terminating each major axis, lateral cymules staminate, axes densely stellate-pubescent; bracts trigonous-subulate, convex, acute, early caducous, densely stellate-tomentose. Flowers unisexual, actinomorphic, fragrant, white to cream; buds ovoid or ovoid-ellipsoid, apex obtuse; calyx valvate, closed in bud, later rupturing to the base into 2 or 3(4) often unequal, lanceolate to ovate lobes, stellate-tomentose outside, subglabrous or with scattered stellate hairs inside; petals 5(6), free, imbricate, oblanceolate-spathulate, rounded at the apex, drying dark brown to black, veins not distinct, outside glabrous or with loose to dense stellate hairs in a longitudinal band in the middle, inside simple-pubescent at the base; disc 5-lobed, lobes free, flattened, inconspicuous. Staminate flowers: pedicel slender, filiform, mostly longer and always significantly thinner than in the pistillate flowers; disc lobes thick, somewhat convolute, glabrous; stamens 17–32, in 4 whorls, the outer ones free, the inner ones united into a column; filaments pubescent, with loose to dense, colourless to brown stellate hairs; anthers basifixed to dorsibasifixed, opening introrsely with 2 longitudinal slits, glabrous or with scattered stellate hairs; pistillode absent. Pistillate flowers: usually somewhat larger than the staminate ones; pedicel short, stout, stellate-pubescent; disc lobes deltoid to semicircular, stellate pubescent, expecially at the apex; ovary superior, 2- or 3(4)-locular, with 1 ovule per locule, densely stellate-tomentose; styles fused at base, thick, bifid, entirely glabrous or stellate-pubescent on the outside. Fruits large, drupaceous, varying in shape according to number of locules developed, evenly to sparingly stellate-pubescent, with fleshy exocarp and thin, woody endocarp. Seeds ecarunculate, subglobose to ovoid, dorsiventrally compressed or not, surface gibberulous; hilum large. Embryo straight, embedded in copious endosperm; cotyledons broad, flat.
Distribution — The genus Aleurites comprises two species. One (A. moluccana) is very widespread (tropical Asia, Oceania from India to China and Polynesia, Australia and New Zealand) and also widely cultivated throughout the tropics, the other (A. rockinghamensis) has only a very limited distribution in Papua New Guinea and Australia.
Note — The name of the genus is derived from the Greek for "wheaten flour", referring to the mealy appearance of the lower leaf surface.
|
1a. |
Inflorescence delicate, base of peduncle 2–4.5(–5) mm diameter. Stamens 17–26. Styles (0.5–)1–2.6 mm long, glabrous or rarely with a few scattered stellate hairs on the outside, bifid almost to the base; ovary 1- or 2(3)-locular. Seeds cordate in dorsal view, dorsiventrally compressed, truncate or rarely minutely tipped at the micropylar end, always tapering towards and pointed at the chalazal end. |
|
|
1b. |
Inflorescence robust, base of peduncle 5–9 mm diameter. Stamens 20–32. Styles 2.6–3.2 mm long, conspicuously stellate-pubescent on the outside, bifid up to half of their length; ovary 3- or 4-locular. Seeds oval to almost elliptic in dorsal view, not dorsiventrally compressed, truncate at the micropylar end, attenuate and obtuse at the chalazal end. |
Aleurites moluccana (L.) Willd., Sp. Pl. 4 (1804) 590; Müll.Arg. in DC., Prodr. 15, 2 (1866) 723; Scheff., Ann. Mus. Bot. Lugduno-Batavum 4 (1868) 120; Hook.f., Fl. Brit. India 5 (1887) 384; Pax in Engl., Pflanzenr. IV.147 (1910) 129, fig. 45; J.J.Sm., Meded. Dept. Landb. Ned.-Indië 10 (1910) 551; Merr., Interpr. Herb. Amboin. (1917) 318; Sp. Blancoan. (1918) 228; J. Straits Branch Roy. Asiat. Soc., special number (1921) 344; Enum. Philipp. Fl. Pl. 2 (1923) 448; Gagnep. in Lecomte, Fl. Indo-Chine 5 (1925) 291; Merr, Trans. Amer. Philos. Soc. new ser., 24 (1935) 239; Backer & Bakh.f., Fl. Java 1 (1963) 478; B.C.Stone, Pacific Sc. 21 (1967) 551; Airy Shaw, Kew Bull. 20 (1967) 393; Kew. Bull. 26 (1972) 213; Whitmore, Tree Fl. Malaya 2 (1973) 54; Airy Shaw, Kew Bull. Add. Ser. 4 (1975) 29; E.Walker, Fl. Okinawa S. Ryukyo Isl. (1976) 644; Airy Shaw, Kew Bull. Add. Ser. 8 (1980) 26; Muelleria 4 (1980) 230; Kew Bull. 36 (1981) 252; Kew Bull. 37 (1982) 5; Alph. Enum. Euphorb. Philipp. Isl. (1983): 4; W.L.Wagner, D.R.Herbst & Sohmer, Man. Fl. Pl. Hawai’i 1 (1990) 598; pl. 80 upper left; P.I.Forst., Muelleria 9 (1996) 7; Stuppy et al., Blumea 44 (1999) 80, Fig. 1j-m; in Chayam. & Welzen, Fl. Thailand 8, 1: 49, fig. 11. 2005. — Jatropha moluccana L., Sp. Pl. (1753) 1006. — Aleurites moluccana (L.) Willd. var. moluccana: Airy Shaw, Muelleria 4 (1980) 230. — (Lecto)type (designated by Stuppy et al., 1999): Hb. Linnaeus 1141/5 (LINN, n.v.; IDC microfiche). See note 1.
Aleurites triloba J.R.Forst. & G.Forst., Char. Gen. Pl. (1776) 56, t. 56.; Boerl., Handl. Fl. Ned. Indië 3, 1 (1900) 282; Hemsl., Bull. Misc. Inform. (1906) 121. — Type: J.R.Forst. & G.Forst., Char. Gen. Pl. (1776) t. 56.
Juglans camirium Lour., Fl. Cochinch. 1 (1790) 573. — Camirium cordifolium Gaertn., Fruct. Sem. Pl. 2 (1791) 195, t. 125, fig. 2. — Aleurites cordifolia Steud., Nomencl. Bot. ed. 2, 1 (1840) 49, nom. superfl. — Type: Rumphius, Herb. Amboin. 2 (1741) t. 58.
Aleurites commutata Geiseler, Croton Monogr. (Mar. 1807) 82. — Type: Vahl s.n. (C, holo, n.v.).
Aleurites ambinux Pers., Syn. Pl. 2 (Sep. 1807) 587. — Lectotype (designated by Stuppy et al., 1999): Anonymous s.n. (P).
[Camirium oleosum Reinw. ex Blume, Catalogus (1823) 104, nom. nud.]
Aleurites lobata Blanco, Fl. Filip. (1837) 756. — Neotype (proposed by Stuppy et al., 1999): Merrill Species Blancoanae 127 (L, holo; iso in NY, W), Philippines, Luzon, Cavite Province, Maragondong.
Aleurites lanceolata Blanco, Fl. Filip. (1837) 757. — Type lost (for synonymy see Fern.-Vill., Nov. App., 1880, 191).
[Aleurites pentaphylla Wall., Numer. List (1847) no. 7959, nom. nud. — pro Wallich 7959 (K, n.v.), India, Calcutta Botanical Garden]
[Aleurites integrifolia Vieill., Ann. Sci. Nat. (Paris) 16 (1862) 59, name in synonymy; Guillaumin, Ann. Inst. Mus. Colon. Marseille, ser. 2, 9 (1911) 225.]
Aleurites angustifolia Vieill., Ann. Sci. Nat. (Paris) 16 (1862) 60; Guillaumin, Ann. Inst. Mus. Colon. Marseille, ser. 2, 9 (1911) 225. — Syntypes: Vieillard s.n. (P?, n.v.), New Caldeonia, Croît à Puébo; Viellard 1149 (P), New Caledonia, Balade.
Aleurites javanica Gand., Bull. Soc. Bot. France 29 (1913) 27. — Type: Treub s.n. (P?, n.v.), Indonesia, Java, cultivated in Bogor Botanical Garden.
Aleurites remyi Sherff, Field Mus. Nat. Hist. Bot. Ser. 17 (1939) 558; Airy Shaw, Kew Bull. 20 (1967) 393. — Aleurites moluccana (L.) Willd. var. remyi (Sherff) B.C.Stone, Pacific Sc. 21 (1967) 553. — Type: Remy 600, p.p. (P, 3 sheets), Hawaii.
Aleurites moluccana (L.) Willd. var. serotina O.Deg. & Sherff, in Sherff, Amer. J. Bot. 38 (1951) 57. — Type: Degener, Clay & Bertram 19329 (F, holo, n.v.; iso in A, B, BISH, CM, G, LA, MO, NY, P, PHIL), U.S.A., Hawaii, Maui, near Kamole Gulch.
Aleurites moluccana (L.) Willd. var. katoi O.Deg., I.Deg. & B.C.Stone [in B.C.Stone, Pacific Sc. 21 (1967) 551, nom. nud.] in O.Deg. & I.Deg., Phytologia 21 (1971) 315. — Type: Stone 3427 (BISH, holo; iso in BISH), U.S.A., Hawaii, Kauai High School in Lihue.
Aleurites moluccana (L.) Willd. var. aulanii O.Deg. & I.Deg., Phytologia 21 (1971) 316. — Type: O. Degener & I. Degener (Stanley & Aulani Loo) 32481 (NY, holo; iso in L), U.S.A., Hawaii, Kukuihaele.
Aleurites erratica O.Deg., I.Deg. & K.Hummel, Phytologia 38 (1978) 361. — Type: O. Degener & I. Degener 24627 (TUB, holo, ?, n.v.; iso in BISH), Phoenix Islands, Canton Atoll.
Croton moluccana auct. non L.: L., Sp. Pl. (1753) 1005 (only reference to Burmann). See note 2.
![]()
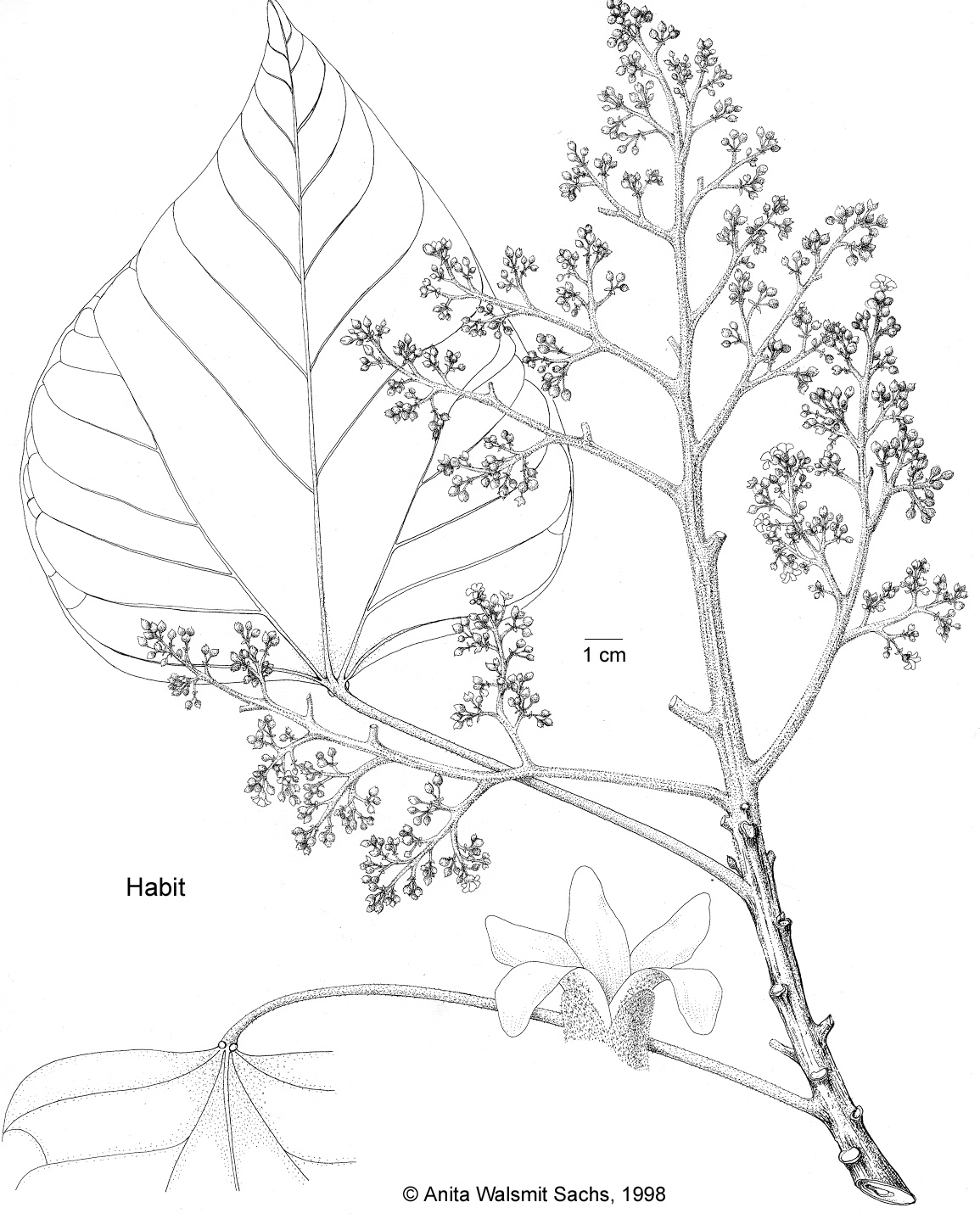
Large tree with spreading pendulous and ascending branches, up to 15–25(–40) m tall. Bark smooth, red-brown, with deep fissures, blaze pink to red. Indumentum greyish- or fulvous-silver to mostly pale, rarely dark ferruginous-silver. Stipules c. 1 mm long. Petiole 3.5–16(–40) cm by 1.5–6 mm; glands at the apex 0.5–2 mm diam. Leaf blade in some extreme, mostly cultivated forms narrowly lanceolate, orbicular or deeply lobed with narrow lobes (see note 3), (6–)12–30 by 3–15(–28) cm, length/width ratio 1–4. Inflorescences delicate, 4–27 by 4–32 cm, axes densely fulvous- (to ferruginous-silver) pubescent; base of peduncle 2–4.5(–5) mm diam.; bracts 2–4 by 0.5–2 mm. Staminate flowers: 5–9 by 5–10 mm; pedicel 4–12 by 0.3–0.5 mm., stellate-pubescent; buds 2–3 by 2–2.5 mm.; calyx lobes 2.5–3 by 1.5–3 mm; petals (3.5–)5–7(–9.5) by 1.2–3 mm; stamens 17–26, outermost filaments 0.5–1 mm long, the innermost ones 2–3.5 mm long; anthers 0.6–0.8 by 0.3–0.6 mm. Pistillate flowers: 6–11 by 7–10 mm; pedicel stout, 2–3.5(–6) by 1–2.5 mm, stellate-pubescent; buds 4–5.5 by 2–2.5 mm; calyx lobes 3–6 by 2–5 mm; petals 6–11 by 1.6–3 mm; ovary subglobose, laterally compressed (rarely trigonous), 2(3)-locular, 1.5–3 by 2–3.5 mm., densely fulvous to rarely ferruginous stellate-tomentose; styles deeply bifid (almost up to the base), (0.5–)1–2.6 mm long, mostly glabrous, rarely with a very few colourless, stellate hairs on the outside. Fruits laterally compressed, broadly ovoid-subglobose or transversely ovoid with 4 (6) low longitudinal ridges, very slightly acuminate at the apex, 4–4.5 by 4–6(–7) cm. Seeds broadly obovoid, cordate in dorsal view, dorsiventrally compressed, truncate or rarely minutely tipped at the micropylar end, always tapering towards the obtusely acuminate chalazal end, (20–)23–32 by 20–32 by (15–)18–24 mm, greyish, mottled brownish.
Distribution — Native of tropical Asia and Oceania, from India and China to Polynesia and New Zealand, and widely cultivated in the tropics generally. According to Forster (1996) in Australia A. moluccana is restricted to north Queensland. About the area of origin of Aleurites moluccana, A.C. Smith (1981) writes: "Indigenous in Malesia, although the precise area of its nativity is probably impossible to establish because of its early aboriginal introduction to neighboring areas, doubtless including India and Ceylon as well as Melanesia and Polynesia."
Habitat & Ecology — Occurring throughout the tropics, wild and cultivated, from sea-level up to at least 1000 m altitude on a variety of substrates, both on red loam and stony clay ground, on sand and limestone, sometimes in periodically rather dry areas (J.J. Smith 1910). Young plants are common as pioneers in disturbed gaps or margins of the vineforest (Forster 1996).
Uses — Wood is used for the production of tea boxes and canoes but not for construction. The seeds are eaten raw or after roasting in fire together with rice or used as purgative. Wrapped in leaves or strung to palm leaf midribs they are also used as torches. The latex is chewed as chewing gum in the Pacific and is also used medicinally and as glue. In Hawaii juice from the husk is used as a black dye, and similarly from the bark of the root, the latter is used to paint canoes. Also in Hawaii, the roasted seeds mixed with salt and seaweed make a condiment called "inamona". Chinese in Hawaii let the wood of felled trees rot to grow a fungus [Auricularia cornea (Ehrenb. ex Fr.) Ehrenb. ex Endl.] for local consumption and for export to China. The tree is also used as an ornamental.
The seeds contain a high amount of oil (60 %) which is used as fuel, lubricant or as part in varnish, paint or soap. The oil is also used as rubbing oil for rheumatism (Pax, 1910; J.J. Smith, 1910; Wagner et al., 1990).
Vernacular names — English: Candle nut, Indian walnut. Indonesia: Kemiri (Malay); moentjang (Sundanese). Philippines: Biau (Bagobo, Cebu Bisaya,); kalumban, kapili, lumbang, lumbang-bato (Tagalog); kami (Sulu); rumbang (Bisaya) (Merrill, 1923). Papua New Guinea: Mboal. Tahiti: Tahii-tairi. Samoa: Lama. Hawaii: Kukui; aulani kukui, small-seeded kukui (former var. alulanii); kato kukui, mango-leaved kukui (former var. katoi); kona kukui (former var. remyi) (Degener & Degener, 1971). Brazil: Noz da India (Pax, 1910).
Notes — 1. Linnaeus in his description refers to his own Ceylon flora and the one made by Hermann. Both refer to the fact that the plant is Moluccan. The specimen in LINN has the reference ‘Comm. Rumphius’ and is one of the Moluccan plants sent by Rumphius (via Hermann?) and may, therefore, be selected as the type.
2. The name Croton moluccana L. is based on two references. The first one (‘Fl. Zeyl. 346’; must be page 146) is Givotia rottleriformis Griff. (Euphorbiaceae), the other one (‘Nux juglans moluccana bifida Burm., Fl. Zeyl. 170’) is Aleurites moluccana (L.) Willd. (the latter is based on Jatropha moluccana L., by coincidence the same epithet). The best thing to do is to lectotypify Linnaeus’s name with one of the Hermann specimens (Givottia rottleriformis), because this is the only indirect reference (via L., Fl. Zeyl.) to specimens; the other reference only mentions the description in Burman’s Thesaurus Zeylanica (= Aleurites moluccana) (Merrill, 1917; Trimen, 1898). The specimen in LINN (no. 1140/20) in London is Melanolepis multiglandulosa, but this is not the type of the species. Müller Argoviensis regarded this specimen as the type and, therefore, erroneously made the necessary combination in Mallotus (and subsequent authors in Rottlera and Melanolepis).
3. As already mentioned in the introduction, Aleurites moluccana shows a strong variation in leaf shape, which has led to the description of several separate species or varieties which are, however, not recognized here. On account of its very narrow, unlobed, lanceolate leaves one New Caledonian form was described as separate species (A. angustifolia) already last century (Vieillard, 1862). Later two extreme forms from Hawaii - according to Stone (1967) probably of aboriginal Hawaiian selection - have been described as separate species or variety respectively, but are not accepted here. One is A. remyi (later assigned to varietal status) bearing relatively lengthened, simple, lanceolate leaves (with or without obscure lobes) or deeply lobed leaves with the lateral lobes very narrow and the terminal lobe much elongated. The second is A. moluccana var. katoi ("Mango-leaved Kukui") bearing narrow, (only?) lanceolate leaves with the lateral lobes obscure or absent (for references see above). Besides that, of the material seen by the authors, one (cultivated) specimen from New Caledonia (Balansa 3443, P) shows perfectly orbicular leaves.
Another dubious variety, A. moluccana var. aulanii ("Small-seeded Kukui"), was described by Degener & Degener (1971) solely on the basis of smaller seeds.
Aleurites rockinghamensis (Baill.) P.I.Forst., Muelleria 9 (1996) 8; Stuppy et al., Blumea 44 (1999) 84, Fig. 1a-i. — Aleurites moluccana (L.) Willd. var. rockinghamensis Baill., Adansonia 6 (1866) 297; Airy Shaw, Muelleria 4 (1980) 230. — Lectotype (designated by Forster, 1996): Dallachy s.n. MEL 232486 (MEL, holo; iso in MEL), Australia, Queensland, Cook District, Rockingham’s Bay.
Aleurites moluccana (L.) Willd. var. floccosa Airy Shaw, Kew Bull. 20 (1966) 26. — Type: NGF (Havel) 9169 (K, holo, n.v.; iso in L, LAE), Papua New Guinea, Morobe Distr., Wau, Anderson’s Logging Area.
![]()
![]()
![]()
Large tree with spreading pendulous and ascending branches, up to 30 m tall. Bark smooth, grey, nondescript, blaze brown speckled to cream. Indumentum sometimes greyish- or fulvous-silver, mostly pale to dark ferruginous-silver. Stipules c. 1 mm long. Petiole 3–21 cm by 2–6 mm; glands at the apex 0.5–2 mm diam. Leaf blade 11–40 by 7–30 cm, length/width ratio 1–1.8. Inflorescences robust, 7.5–38 by 7.5–30 cm, axes densely fulvous- to ferruginous-silver stellate-pubescent; base of peduncle (5–)6 mm diam.; bracts 3–5 by 0.8–2 mm. Staminate flowers 5–12 by 6–12 mm; pedicel 5–15 by 0.3–0.6 mm, stellate-pubescent; buds 3.5–5 by 3–4 mm; calyx lobes 3–5.8 by 2.5–5 mm; petals 4.5–10 by 2–4 mm; stamens 20–32, outermost filaments 0.5–1 mm long, the innermost ones 2–3.8 mm long; anthers 0.6–0.9 by 0.3–0.7 mm. Pistillate flowers: 8–12 by 8–12 mm; pedicel stout, 2–5.5 by 2–3 mm, stellate-pubescent; buds 4.5–6 by 3–3.5 mm; calyx lobes 4–7 by 3–6 mm; petals 9–13.5 by 2.5–4 mm; ovary subglobose, trigonous (or rarely tetragonous) in cross-section, 3(4)-locular, 2–3 by 2.5–3.5 mm, densely fulvous to ferruginous stellate-tomentose; styles bifid up to half of their length, 2.6–3.2 mm long, densely colourless to ferruginous stellate-pubescent without, glabrous within. Fruits broadly ovoid-subglobose with 6(8) low longitudinal ridges, very slightly acuminate at the apex, 4–6.5 by 5.5–8 cm. Seeds subglobose to obovoid, oval to almost elliptic in dorsal view, not dorsiventrally compressed, truncate at the micropylar end, attenuate and obtuse at the chalazal end, 20–27 by 20–25 by 20–25 mm, white to cream, conspicuously mottled brown.
Distribution — Australia (Queensland with a disjunct southerly occurrence near Ingham; Forster, 1996) and Papua New Guinea.
Habitat & Ecology — Rather common tree in rain forest, also occurring in the margins of the forests and as a pioneer in disturbed places, thriving on a variety of substrates usually of volcanic origin, up to at least 1190 m altitude.
Vernacular names — English: Candle nut. New Guinea: Tsikúreh (Managalase), Towai (Daga, Bonenau).
Uses — Endosperm edible after cooking or roasting.