
|
Malesian Euphorbiaceae Descriptions |
|
Esser, H.-J. 1999. A partial revision of the Hippomaneae (Euphorbiaceae) in Malesia. Blumea 44: 149–215.
Goto on this page:
Microstachys A.Juss., Euph. Gen. (Feb. 1824) 48, t. 15, fig. 50; Müll.Arg., Linnaea 32 (1863) 90; G.L.Webster, Ann. Missouri Bot. Gard. 81 (1994) 122; Esser, Kew Bull. 53 (1998) 955; Blumea 44 (1999) 175; in Radcl.-Sm., Gen. Euphorbiacearum (2001) 365; Esser & Welzen in Welzen & Chayam., Fl. Thailand 8, 2 (2007) 452; G.L.Webster in Kubitzki, Fam. Gen. Vasc. Pl. 11 (2014) 196, Fig. 43. — Sebastiania sect. Microstachys (A.Juss.) Müll.Arg. in DC., Prodr. 15, 2 (1866) 1166; Benth. in Benth. & Hook.f., Gen. Pl. 3 (1880) 336; Pax in Engl. & Prantl, Nat. Pflanzenfam. 3, 5 (1890) 94; Pax & K.Hoffm. in Engl., Pflanzenr. IV.147.v (1912) 91; in Engl. & Harms, Nat. Pflanzenfam. ed.2, 19c (1931) 192. — Sapium sect. Sebastiania (Spreng.) T.Post & Kuntze subsect. Microstachys (A.Juss.) T.Post & Kuntze, Lex. Gen. Phan. (1903) 498. — Lectotype (designated by G.L.Webster, Ann. Missouri Bot. Gard. 81, 1994: 121): Microstachys bicornis (Vahl) A.Juss., based on Tragia bicornis Vahl [= Microstachys corniculata (Vahl) Griseb.].
Cnemidostachys Mart. & Zucc., Flora 7.1 Beibl. 4 (Apr.–June 1824) 136; Nov. Gen. Sp. Pl. 1 (Oct. 1824) 66, t. 40. — Stillingia sect. Cnemidostachys (Mart. & Zucc.) Baill., Étude Euphorb. (1858) 515; Adansonia 1 (1861) 350. — Excoecaria sect. Cnemidostachys (Mart. & Zucc.) Baill., Adansonia 6 (1867) 323. — Sebastiania sect. Cnemidostachys (Mart. & Zucc.) G.L.Webster, J. Arnold Arbor. 48 (1967) 386. — Lectotype (designated by Esser, 1999): Cnemidostachys myrtilloides Mart. & Zucc. [= Microstachys daphnoides (Mart. & Zucc.) Müll. Arg.]; see note 2.
Elachocroton F.Muell., Hook. J. Bot. Kew Gard. Misc. 9 (1857) 17. — Stillingia sect. Elachocroton (F.Muell.) Baill., Étude Euphorb. (1858) 517. — Sebastiania sect. Elachocroton (F.Muell.) Pax in Engl., Pflanzenr. IV.147.v (1912) 114; Pax & K.Hoffm. in Engl. & Harms, Nat. Pflanzenfam. ed. 2, 19c (1931) 192; A.S. de Oliveira, Arq. Jard. Bot. Rio de Janeiro 27 (1983) 3. — Type: Elachocroton asperococcus F.Muell. [= Microstachys chamaelea (L.) Müll. Arg.].
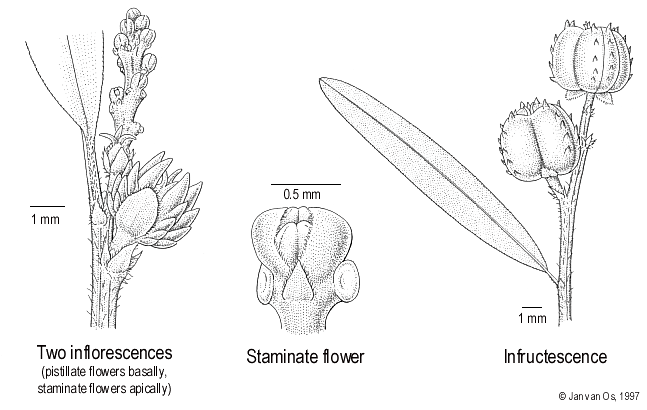
Herbs to subshrubs, less than 2 m high. Monoecious. Flowering and fruiting twigs with leaves. Indumentum consisting of multicellular, uniseriate, colourless hairs, mostly on vegetative parts, rarely absent. Stipules triangular, 0.5–2 mm long, undivided or apically divided into several ciliae, glandless. Leaves regularly alternate; petiole short (less than 1 cm long; absent to long in some Neotropical species), glandless; blade elliptic, ovate or linear, 0.3–2 cm wide, base acute to subcordate, margin with very dense, minute and persistent glandular teeth, 0.3–0.6 mm apart and often nearly touching each other (rarely fused into an entire, glandulous margin in some Neotropical taxa), apex rounded, acute or mucronate, above glandless and glabrous, lower surface paler and smooth to papillate but never white, glabrous to pilose, at base often with few marginal to submarginal glands, basal ones slightly larger, secondary veins arching and usually looped but often hardly visible, tertiary veins not visible but presumably reticulate. Inflorescences terminal and axillary, often opposite to leaves, yellowish, simple, without sterile basal region, pistillate and staminate flowers in same thyrse or pistillate flowers separated, staminate thyrse 5–60 by 1–2 mm, glabrous to pilose. Bracts of staminate cymules triangular, acute, with a pair of flattened-orbicular to falcate glands touching the axis of the thyrse. Staminate cymules 1–3-flowered; bracteoles absent. Staminate flowers with very short (less than 0.5 mm long) pedicel, hardly elongating when flowering; calyx with 3 spathulate, quite large, free sepals; stamens 3, filaments longer than anthers. Pistillate flowers 1 (very rarely 2) at base of staminate thyrse or often separated from staminate part and on different node, sessile; calyx with 3 sepals, triangular, free, glandless or with a pair of minute basal glands; ovary 3-locular, usually with 2 longitudinal rows of spine-like excrescences on each carpel, glabrous to hirsute; style usually absent, stigmata 3, undivided, glandless. Fruits with short (0–2 mm long) pedicel; 3-seeded, oblong with very regular shape, usually with 6 rows of spine-like excrescences, dry, dehiscing regularly along the septa, glabrous to hirsute; mericarps with a thin exocarp (fruit length/pericarp thickness > 10/1), septa very regular, with a small separate basal triangle and 1 furcate vascular strand; remaining central columella very regular, slightly alate with parallel margins over its whole length. Seeds elliptic-oblong with truncate ends, dry, with a large persistent caruncle stipitate and lenticular when dry.
Distribution — 15 species currently accepted, 10 of them in tropical South America south of the Amazon, one in northern South America and Central America up to Mexico, three endemic to Africa, and one in the Old World from Africa through Asia to N Australia (absent from the Philippines and not yet known from New Guinea).
Notes — 1. In the New World, the distinction between the species has notoriously posed problems, based primarily on characters of indumentum and leaf shape. The distinctiveness of the only Asian species, however, has never been questioned.
2. Von Martius & Zuccarini described 17 species of Cnemidostachys simultaneously, and several previous authors avoided to select a lectotype (e.g. Webster, 1994). However, the plate illustrating C. myrtilloides is also cited as the one accompanying the diagnosis of the genus, and was obviously meant as illustration representative for the genus. For this reason C. myrtilloides is proposed here as lectotype.
Microstachys chamaelea (L.) Müll.Arg.
Microstachys chamaelea (L.) Müll.Arg., Linnaea 32 (1863) 95, see note 1; Esser, Blumea 44 (1999) 176, Fig 4, Map 4; Esser & Welzen in Welzen & Chayam., Fl. Thailand 8, 2 (2007) 453, Fig. 46, Plate XXII: 3. — Tragia chamaelea L., Sp. Pl. (1753) 981. — Cnemidostachys chamaelea (L.) Spreng., Syst. Veg. 3 (1826) 835. — Excoecaria chamaelea (L.) Baill., Adansonia 6 (1867) 323. — Sebastiania chamaelea (L.) Müll.Arg. in DC., Prodr. 15, 2 (1866) 1175; Benth., Fl. Austral. 6 (1873) 151; Hook.f., Fl. Brit. India 5 (1888) 475; Kuntze, Revis. Gen. Pl. 2 (1891) 619; Boerl., Handl. Fl. Ned. Ind. 3, 1 (1900) 268, 296; Pax & K.Hoffm. in Engl., Pflanzenr IV.147.v (1912) 116; Merr., Philipp. J. Sc. 10, C Bot. (1915) 190; Ridl., Fl. Malay Penins. 3 (1924) 317; Gagnep. in Lecomte, Fl. Indo-Chine 5 (1926) 454, fig. 55.5–14; Burkill, Dict. Econ. Prod. Malay Pen. 2 (1935) 1988; M.R.Hend., Malay. Wild Fl. (1959) 464, fig. 418; Nair & Maitreyi, Bot. Gaz. 124 (1962) 58; Backer & Bakh.f., Fl. Java 1 (1964) 498; Airy Shaw, Kew Bull. 26 (1972) 339; Whitmore, Tree Fl. Malaya 2 (1973) 131; Airy Shaw, Kew Bull. Add. Ser. 4 (1975) 195; Kew Bull. 35 (1980) 686; Muelleria 4 (1980) 244; H.Keng, Conc. Fl. Sing. (1990) 113; P.H.Hô, Câyco Viêtnam 2, 1 (1992) 352; I.M.Turner, Gard. Bull. 45 (1993) 87; Gard. Bull. 47 (1995) 232; Y.C.Tseng, Fl. Reipubl. Pop. Sin. 44(3) (1997) 4, pl. 1. — Lectotype (designated by Esser, 1999): Herb. Hermann vol. 4: 43, no. 335 (BM), Ceylon.
Elachocroton asperococcus F.Muell., Hook. J. Bot. Kew Gard. Misc. 9 (1857) 17. — Sebastiania chamaelea (L.) Müll.Arg. var. asperococca (F.Muell.) Pax in Engl., Pflanzenr. IV.147.v (1912) 117; Merr., Philipp. J. Sc. 11, C Bot. (1916) 76; Pax & K.Hoffm. in Engl., Pflanzenr. IV.147.xiv, add. VI (1919) 58; Merr., Bibl. Enum. Bornean Pl. (1921) 347. — Type: F. Müller s.n. (holo MEL, n.v.; iso K), Australia, ”in locis sterilioribus ad flumen Victoriae”.
Cnemidostachys linearifolia Miq., Fl. Ind. Bat. Suppl. 1 (1861) 460. — Type: Kurz (as "Amann") 9 (holo U; iso K), Java, Banka, "in pratis aridis ad rivulam Muntok".
![]()
![]()
Perennial herb to many-stemmed subshrub, up to 90 cm tall and 30 cm in total diameter; originating from a yellowish-brown tap root c. 15 cm long. Indumentum 0.25–0.4 mm long. Stipules 0.6–2 by 0.2–0.5 mm, densely pilose to ciliate. Leaves: petiole 0.1–0.25 cm long, pilose, glabrescent; blade elliptic to lineal, 1.6–6 by 0.3–0.8(–1.2) cm, base acute to obtuse, margin with ± appressed teeth, apex rounded to mucronate, lower surface bright but smooth, pilose and glabrescent, with 0–6 strictly marginal glands per side, 0.2–0.25 mm in diam., secondary veins 13–35 pairs, angle with midrib 45–75°, often hardly visible. Inflorescences: staminate part 5–15 by 1–1.5 mm, axis usually glabrous. Bracts of staminate cymules c. 0.4 mm long, nearly completely covered by the bract glands. Staminate cymules 1-flowered. Staminate flowers nearly sessile (pedicel 0.2–0.4 mm long); calyx 0.5 mm long, with distinctly ciliate sepals. Pistillate flowers: calyx 0.6–1.2 mm long, with distinctly ciliate sepals; ovary glabrous; stigmata filiform, c. 0.5–1 by 0.1 mm. Fruits 3.5–4.5 mm long, green, turning pinkish to olive brown to orange, with 0.6–1 mm long excrescences in 6 rows of 5–6 each; pericarp c. 0.15 mm thick. Seeds 2.5–2.75 by 2–2.25 mm, brown, sometimes brown-yellowish-whitish mottled.
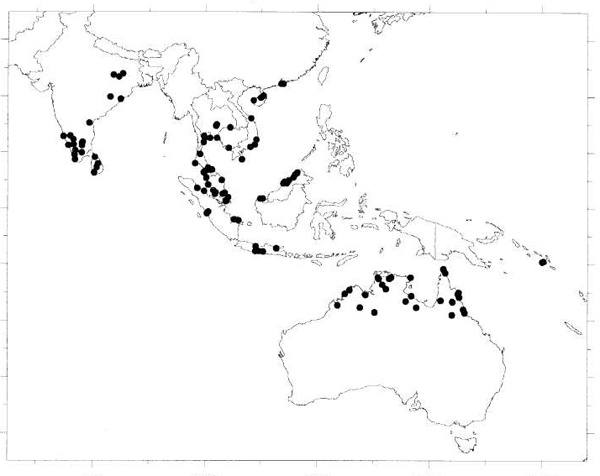
Distribution — Widespread in Africa (e.g., Camerun, Togo) and in Asia from India and Sri Lanka to Vietnam and S China, the Solomon Islands and N Australia; Malesia: Malay Peninsula (Thailand, Malaya, Singapore), Sumatra, Java, Borneo (Brunei, Sabah, Sarawak).
Habitat & Ecology — Found on beaches, sandy sites, roadsides and waste places, also on lateritic outcrops, dry dipterocarp forest, grassfields, and open woodland, often gregarious. Soil: sand, alluvial and lateritic ground. Altitude sea level up to 300 m. Flowering and fruiting the whole year through, but particularly in the rainy season.
Uses — When cooked together with meat and vegetables, whole young plants are used for speedy recovery for women after giving birth [S (Mamit) 35141]. Dragendorff (1898; repeated by Burkill, 1935) mentions the use of the plant as adstringent, tonic, against diarrhoea and syphilis in India.
Vernacular names — Malay Peninsula: Ambin-ambin (Burkill, 1935). Borneo: Daun merayat (Lundu Malay).
Notes — 1. De Jussieu (1824), when describing Microstachys, mentioned Tragia chamaelea as belonging to his new genus. But because he did not definitely combine genus and species, the combination Microstachys chamaelea cannot be attributed to him (Art. 33.1 ICBN). Baillon (1858) cited this combination only as a synonym, which also does not constitue a valid publication (Art. 34.1 ICBN). Müller Argoviensis (1863), therefore, was the first to use and accept this name.
2. There has been confusion in the labels and the literature between Cuming 2324 and Cuming 2329. Müller Argoviensis (1866) and Pax & Hoffmann (1912) cited both specimens under Sebastiania (Microstachys) chamaelea. Nevertheless, Cuming 2324 (K) is the type collection of Sarcotheca monophylla (Planch. ex Hook. f.) Hallier f., based on Connaropsis monophylla Planch. ex Hook. f., an Oxalidacea. At K (including the herbaria of Bentham and Hooker), only Cuming 2329 can be referred to M. chamaelea; the same is true for FR. At L, however, only Cuming 2324 is an originally labelled M. chamaelea, and Cuming 2329 was not found at all. Thanks are due to J.F. Veldkamp who recognized this confusion. Both specimens, labelled as collected on the Philippines, really are from Malaya.
3. Contrary to data in the literature, leaves with entire margins were never found, although the appressed teeth are very small and may be overlooked. In Malesia, the appendages of the fruits are a very regular feature. Those of Australian plants, however, are weaker to totally absent. Specimens from Australia may also be distinguished by their often wider leaves.
4. Microstachys chamaelea is sometimes confused with Sauropus bacciformis (L.) Airy Shaw in the herbaria, a herb with similar habit but entire leaves and smooth fruits with petals and carinate, reddish seeds.