
|
Malesian Euphorbiaceae Descriptions |
|
Esser, H.-J. 1997. A revision of Omalanthus (Euphorbiaceae) in Malesia. Blumea 42: 421–466.
Goto on this page:
Key to the species using Leaves and Staminate Inflorescences
Key to the species using Leaves and Fruits
(H)Omalanthus A.Juss., Euphorb. Gen. (1824) 50, t. 16 fig. 53; Blume, Bijdr. 12 (1825) 627; Endl., Gen. Pl. (1840) 1110; Baill., Étude Euphorb. (1858) 537, Atlas pl. 8, fig. 22–31; Kuntze, Revis. Gen. Pl. 2 (1891) 609; G.L.Webster, Taxon 24 (1975) 600; St. John, Nord. J. Bot. 4 (1984) 53; G.L.Webster, Ann. Missouri Bot. Gard. 81 (1994) 120; Esser, Blumea 42 (1997) 427; in Radcl.-Sm., Gen. Euphorbiacearum (2001) 354; Esser & Welzen in Welzen & Chayam., Fl. Thailand 8, 2 (2007) 334; G.L.Webster in Kubitzki, Fam. Gen. Vasc. Pl. 11 (2014) 196. — Homalanthus Rchb., Consp. Regn. Veg. (1828) 194; Hassk., Cat. Hort. Bot. Bogor. (1844) 233; Benth., J. Linn. Soc., Bot. 17 (1878) 194, 240; in Benth. & Hook.f., Gen. Pl. 3.1 (1880) 331; Pax in Engl. & Prantl, Nat. Pflanzenfam. 3.5 (1890) 95; J.J.Sm. in Koord. & Valeton, Bijdr. Boomsoort. Java 12 (1910) 619; Pax & K.Hoffm. in Engl., Pflanzenr. IV, 147, v (1912) 42; in Engl. & Harms, Nat. Pflanzenfam. ed. 2, 19c (1931) 188; Chittenden, Dict. Gard. 2 (1951) 1006; Airy Shaw, Kew Bull. Add. Ser. 8 (1980) 115; Huxley, New R.H.S. Dict. Gard. 2 (1992) 578. — Homalanthus Sect. Euhomalanthus Kuntze in T.Post & Kuntze, Lex. Gen. Phan. (1903) 285, nom. inval. — Type: Homalanthus leschenaultianus A.Juss. [= Homalanthus populneus (Geiseler) Pax].
Duania Noronha, Verh. Batav. Genootsch. Kunst Wet. 5 ed. 1 (1790) 2 (nomen); ed. 2 (1827) 65 (nomen).
Carumbium Reinw., Elench. Sem. Hort. Leiden ex Isis 1 (1823) 319 (nomen); in Blume, Cat. Pl. Bogor. (1823) 105 (nomen); Syll. Pl. Nov. 2 (1825) 6; Baill., Adansonia 6 (1867) 348; Benth., Fl. Austr. 6 (1873) 149. — Carumbium sect. Eucarumbium Müll.Arg. in DC., Prodr. 15.2 (1866) 1143, nom. inval. — Type: Carumbium populifolium Reinw. [= Homalanthus populneus (Geiseler) Pax].
Dibrachion Regel, Index Sem. Hortus Bot. Petropol. 1865 (1866) 51 (nomen illeg.; see note 1); Gartenflora 15 (1866) 100, Tafel 504. — Type: Dibrachion peltatum Regel [= Homalanthus fastuosus (Linden) Fern.-Vill.].
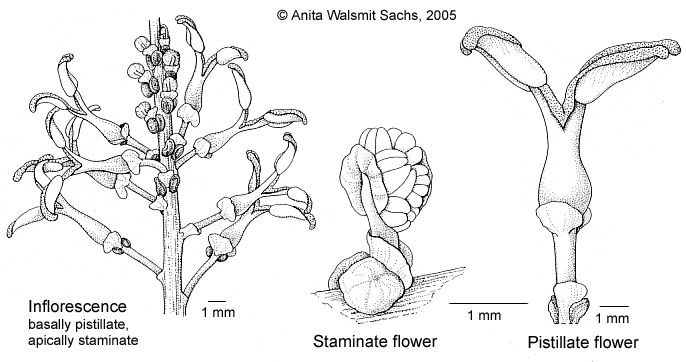
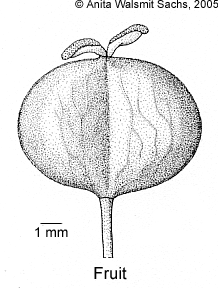
Trees, with pseudoverticillate branching. Monoecious. Bark without spines, nearly smooth. Latex in all parts, white, sticky. Wood white to straw, soft. Indumentum consisting of uniseriate, multicellular, pale to brownish hairs, but absent in most species. Stipules remarkably large (0.5–12 cm long), cuneate at base, margin entire, truncate to acute at apex, with distinct parallel venation, enclosing buds of leaves and inflorescences, usually present on youngest shoots but absent on older ones. Leaves alternate, simple; petiole present and shorter to or nearly as long as lamina, adaxially canaliculate, glandless or apically with 1 or 2 adaxial glands, 2 abaxial glands or a cluster of glands surrounding the apex; lamina symmetric, base cuneate to emarginate, not to distinctly peltate, very base often attenuate, margin entire (outside of Malesia rarely remotely crenate), flat, glandless, apex rounded to cuspidate, lower surface usually whitish, sometimes only between veins, sometimes on nearly the whole surface, side veins in 8–15 pairs below the apex, looped, basal one not different, tertiary veins always distinct, numerous, percurrent to (rarely) reticulate, quarternary veins distinct to indistinct, reticulate to percurrent, adaxially glandless or with glandular margin at base or with 1 or 2 cup- to disc-shaped, prominent glands near the junction with the petiole, abaxial glands present in all species, absent only on single specimens, in 1–2 submarginal to laminar rows close to the margin (then near side veins in axils of divisions) or close to midrib, basal pair often distinctly enlarged and remote from to touching petiole or even on abaxial apex of petiole. Inflorescences thyrsal, with an unbranched axis (not compound), bisexual, on single specimens sometimes only with one sex, with spirally arranged bracts, basal ones with a pistillate or, rarely, with a pistillate and few staminate flowers, distal ones with cymules of staminate flowers. Bracts of staminate cymules persistent, triangular to ovate, margin ± entire, apex acute to truncate, at marginal base with one to several pairs of glands, this pair sometime situated even dorsally or on the thyrsal axis. Staminate flowers 1–3(–8) per cymule, inclinate in bud, free, yellowish, pedicel present, articulate; calyx large, persistent, bilateral to inclinate, consisting of 1 or 2 kidney-shaped, entire, free lobes; corolla, disc, and nectaries absent; stamens 4–c.30 per flower, free, usually covered with dense papillae, filaments present, nearly as long as anthers, hardly elongating at anthesis, anthers dorsifix, opening latro-extrorsely with longitudinal slits, connective present; pistillode absent. Pistillate flowers inclinate in bud; bract caducous, similar to but larger than that of staminate cymules; pedicel present, not articulate, elongating in fruit; calyx persistent to caducous, consisting of 1–3 kidney-shaped, entire, free lobes, apex obtuse to truncate, glandless or with basimarginal glands; corolla, disc, and staminodes absent; ovary 2- or 3-locular, without excrescences, mostly papillate to puberulous, each locule with 1 anatropous ovule, style very short to distinct, persistent (early caducous only in H. macradenius), stigma longer than wide, usually recurved, undivided or divided apically to over its whole length, in all species with an apical to subapical gland beneath (in few species often absent). Fruit a woody schizocarp, 2- or 3-seeded, sometimes with only 1 seed developed and then with lateral style, opening primarily loculicidally, pericarp thick and then opening regularly, or pericarp thin and then opening tardily to irregularly; remaining columella slightly alate. Seeds basically elliptic but usually irregularly foveolate and often flattened, testa reddish to brownish, smooth, not shining, only apically or over its whole length covered by pale to whitish, membranaceous arillode.
Distribution — C. 22 species, distributed from the Pacific islands to New Caledonia, Australia, and throughout Malesia including the southern border of Thailand and the Orchid Island of Taiwan. Most Malesian species are endemic to Malesia; only two (H. novoguineënsis, H. populifolius) are also distributed in Australia (both) and New Zealand (H. populifolius). Additionally, H. populifolius is often planted in botanical gardens; the occurrence in Sri Lanka probably also originates from naturalized bed plants.
Notes — 1. The name Dibrachion is a later homonym of Dibrachion Tulasne, a legume genus described in 1843.
2. In the following, the different spellings of Omalanthus vs. Homalanthus are not mentioned any more. The usage by different authors may be inferred from the heading of the genus.
|
1a. |
Leafblade whitish beneath with abaxially very distinct, completely visible venation; abaxial laminar glands close to midvein. Petiolar glands present, lateral to adaxial.— Borneo. |
|
|
1b. |
Leafblade whitish beneath or not, with either abaxially completely visible venation, or only larger veins distinct; abaxial laminar glands, if present, between midvein and margin or closer to margin. Petiolar glands absent or lateral to abaxial (rarely partially adaxial in H. giganteus). |
|
|
2a. |
Leaves with abaxial to lateral petiolar glands. |
|
|
2b. |
Leaves without abaxial to lateral petiolar glands. |
|
|
3a. |
Staminate flowers 1 per bract, with one calyx lobe and (6–)12–30 stamens. Petiolar glands sometimes irregularly enlarged or multiple. |
|
|
3b. |
Staminate flowers 1–3 per bract, with two calyx lobes and 6–10(–12) stamens. Petiolar glands always in one pair and regularly disc-shaped. |
|
|
4a. |
Bract glands 0.25–0.75 mm long, distinctly shorter than the bract. Leaves apically mucronate to acuminate, often yellowish villose at base. Petiolar glands not enlarged but often in several pairs. |
|
|
4b. |
Bract glands 1–1.5 mm long, compeletely covering the bract. Leaves apically rounded to acute to mucronate, always glabrous. Petiolar glands often enlarged and irregularly foliaceous. |
|
|
5a. |
Bract glands 1–2 mm long, nearly as long as bract.— Philippines. |
|
|
5b. |
Bract glands 0.5–1.25 mm long, distinctly shorter than bract. — New Guinea, Solomon Islands. |
|
|
6a. |
Staminate flowers (1–)3 per bract, with 6–8(–10) stamens.— New Guinea. |
|
|
6b. |
Staminate flowers 1 per bract, with (6–)9–12 stamens. — Solomon Islands. |
|
|
7a. |
Leaves on the upper side with one (or two) prominent, cup- to disc-shaped glands at junction lamina base/petiole apex. |
|
|
7b. |
Leaves on the upper side at junction lamina base/petiole apex without a prominent gland, sometimes with inconspicuous glandular tissue or 2 small glands. |
|
|
8a. |
Bract glands elevated by peduncle, not touching rachis. |
|
|
8b. |
Bract glands not elevated, touching rachis. |
|
|
9a. |
Bract glands multiple, divided into several pairs. |
|
|
9b. |
Bract glands simple, only in 1 pair. |
|
|
10a. |
Bract glands in 1–3 pairs, each gland pillow-shaped and uniformly glaucous-papillate. Plant glabrous. |
|
|
10b. |
Bract glands in many, at least 4, pairs, each gland cup-shaped with shiny center and paler, glaucous-papillate, usually elevated margin. Plant glabrous or puberulous. |
|
|
11a. |
Leafblade narrowly elliptic, index 1.8–2.5(–4.6) 12 |
|
|
11b. |
Leafblade orbiculate to elliptic, index 1.1–1.8 |
|
|
12a. |
Leafblade distinctly whitish beneath, usually with reticulate tertiary veins and without conspicuous basilaminar glands. |
|
|
12b. |
Leafblade usually not whitish but shiny beneath, with percurrent tertiary veins and with conspicuous, 0.75–3 mm long, basilaminar glands. |
|
|
13a. |
Leafblade distinctly whitish beneath, with small or without basilaminar glands. Bract glands only 0.25 (–0.6) mm long, glaucous-papillate, often absent on some bracts. |
|
|
13b. |
Leafblade usually not whitish but shiny beneath, with conspicuous, 0.75–3 mm long, basilaminar glands. Bract glands 0.6–1 mm long, shiny, present on all bracts. |
|
|
14a. |
Leafblade narrowly elliptic (index 1.8–4.6), not peltate, with usually reticulate tertiary venation, without conspicuous abaxial, basilaminar glands. Staminate cymules 1(–3)-flowered. Bract glands 0.2–0.25(–0.5) mm long, distinctly (at least 0.5 mm) overtopped by their bracts. |
|
|
14b. |
Leafblade orbiculate to narrowly elliptic (index 0.8–4.6), peltate or not, with percurrent tertiary venation, with or without conspicuous abaxial, basilaminar glands. Staminate cymules (1–)3-flowered. Bract glands 0.5–1.5 mm long, hardly (up to 0.3 mm) overtopped by their bracts. |
|
|
15a. |
Leafblade not peltate, with 1–3 pairs of abaxial, laminar glands. Usually in the lowland, rarely above 1,400 m elevation. |
|
|
15b. |
Leafblade peltate or not, with 5–28 pairs of abaxial, laminar glands. Only known from above 1,400 m elevation. |
|
1a. |
Fruits regularly trilocular. — Solomon Islands. |
|
|
1b. |
Fruits uni- to bilocular, rarely single ones trilocular. |
|
|
Style and stigma very soon caducous, aborted at base and never found on ripe fruits. Fruits very small (3.5–4.5 mm long) and deeply sulcate. Leaves apically mucronate to acute to rounded; petiolar glands often enlarged to up to 6 mm long, irregular appendages, rarely regularly disc-shaped. |
||
|
2b. |
Style and stigma not very soon caducous, often found on ripe fruits. Fruits (3.5–)5–14 mm long, sulcate or not. Leaves apically mucronate to cuspidate; petiolar glands absent or regularly rounded, disc- or cup-shaped. |
|
|
Stigma glandular over nearly its whole length, only up to 0.5 mm glandless. Leaves without conspicuous and prominent, adaxial or petiolar glands. |
||
|
3b. |
Stigma not glandular over its whole length, more than 0.5 mm glandless. Leaves with or without conspicuous and prominent, adaxial or petiolar glands. |
|
|
Stigma usually distinctly divided, often over its whole length (then apparently 4 per fruit), only rarely undivided. Fruit usually club-shaped with a 1–3 mm long, narrow base. Leaves with simple or multiple petiolar glands, often with distinct yellowish brown, villose indumentum. |
||
|
4b. |
Stigma not or, rarely, only apically divided. Fruit elliptical to slightly obovoid in shape, without a conspicuous, narrow base. Leaves without or with simple petiolar glands, glabrous or hairy. |
|
|
Leaf blades with very distinct, abaxially completely visible venation, and abaxial laminar glands close to the midvein. — Borneo. |
||
|
5b. |
Leaf blades with abaxially completely or, more often, only partially visible venation, and with laminar glands absent, between midvein and margin, or closer to the margin. |
|
|
Leaves adaxially with one or two elevated glands at junction blade/petiole. |
||
|
6b. |
Leaves adaxially glandless or with two inconspicuous often diffuse glands. |
|
|
Stigmatic glands covering approximately half of the length of the stigma. |
||
|
7b. |
Stigmatic glands very short, covering less than a quarter of the length of the stigma, or absent. |
|
|
Leaf blades abaxially usually not whitish, rarely whitish. 2–7 fruits per infructescence. |
||
|
8b. |
Leaf blades whitish beneath. 1 fruit per infructescence. |
|
|
Leafblade narrowly elliptic, index 1.8–4.6. Stigma often widest at base and continuously narrowing. |
||
|
9b. |
Leafblade narrowly or broadly elliptic, index 0.8–1.8. Stigma usually not conspicuously wider at base. |
|
|
Bract glands in 1–3 pairs, each gland pillow-shaped, uniformly glaucous-papillate without conspicuous margin. Fruits 1–4 per infructescence. Plant glabrous. |
||
|
10b. |
Bract glands in at least 4 pairs, each gland cup-shaped with shiny center and thickened, glaucous-papillate margin. Fruits 2–30 per infructescence. Plant glabrous or puberulous. |
|
|
Leaves with large (1–2 mm long) glands on the abaxial lamina base close to the petiole, but without true petiolar glands.— W Malesia. |
||
|
11b. |
Leaves with smaller (0.25–0.75 mm long) glands on the abaxial lamina base, with or without true petiolar glands. |
|
|
Leaves without abaxial-lateral petiolar glands. |
||
|
12b. |
Leaves with abaxial-lateral petiolar glands visible from beneath. — E and C Malesia. |
|
|
Fruits often with lateral hooks giving an obovoid shape; bracts totally covered by bract glands. — Philippines. |
||
|
13b. |
Fruits orbicular in shape, without lateral hooks; bracts distinctly longer than their glands. — New Guinea. |
Homalanthus arfakiensis Hutch. in Gibbs, Fl. Arfak Mts. (1917) 145; Pax & K.Hoffm. in Engl., Pflanzenr. IV, 147, xvii, add. VII (1924) 198; Airy Shaw, Kew Bull. 21 (1968) 415; Kew Bull. Add. Ser. 8 (1980) 117; P. van Royen, Alpine Fl. New Guinea 3 (1982) 2179, fig. 662; Esser, Blumea 42 (1997) 431, Map 1. — Type: Gibbs 5966 (BM holo; iso K), New Guinea, Irian Jaya, Arfak Mts., Anggi lakes.
Homalanthus collinus Gage in Herderschee, Nova Guinea 12 (1917) 484; Pax & K.Hoffm. in Engl., Pflanzenr. IV, 147, xvii, add. VII (1924) 199; Airy Shaw, Kew Bull. 21 (1968) 415; Kew Bull. Add. Ser. 8 (1980) 117. — Type: Pulle 1013 (BO holo, n.v.; iso K, L, U), New Guinea, Irian Jaya, summit of Mount Wichmann.
Homalanthus megalanthus Gage in Herderschee, Nova Guinea 12 (1917) 482, t. 184; Pax & K.Hoffm. in Engl., Pflanzenr. IV, 147, xvii, add. VII (1924) 199; Airy Shaw, Kew Bull. 21 (1968) 415; Kew Bull. Add. Ser. 8 (1980) 117. — Type: Pulle 2409, leg. Versteeg (BO holo, n.v.; iso K, L, U), New Guinea, Irian Jaya, Mount Hubrecht.
Homalanthus agallochoides J.J.Sm. in Herderschee, Nova Guinea 12 (1917) 547, pl. 228B, 229; Pax & K.Hoffm. in Engl., Pflanzenr. IV, 147, xvii, add. VII (1924) 199; Airy Shaw, Kew Bull. 21 (1968) 415. — Type: Gjellerup 1206 (BO holo, n.v.; iso L), New Guinea, Irian Jaya, Arfak-Gebirge, zwischen den Anggi-Seen.
?Homalanthus minutiflorus Airy Shaw, Kew Bull. 21 (1968) 414; Kew Bull. 35 (1980) 399; Kew Bull. 37 (1982) 25. — Type: Kostermans 884 (K holo; iso L), Moluccas, Morotai, Sambiki River.
Tree up to 40 m tall, dbh up to 25 cm, widely spreading and densely foliaged. Glabrous. Bark c. 3 mm thick; outer one medium brown, with frequent horizontal, large, white lenticels; inner one pale green to yellowish straw. Wood with purple discoloration. Stipules 1.5–3 cm long. Leaves: petiole 0.8–5 cm long, glandless; lamina elliptic to ovate, 3–11 by 2–5 cm, index 1.8–2.5(–4.6), base attenuate to obtuse to rounded, not peltate, margin often reddish, apex acute to acuminate, lower surface whitish but most of venation of different colour, side veins in 8–14 pairs below the apex, angle of divergence 50–55°, hardly joined towards the margin, tertiary veins indistinctly percurrent to reticulate, quarternary veins reticulate, adaxially usually glandless or, rarely, with a pair of disc-shaped and prominent glands on junction lamina base/petiole, each gland 0.75–1.25 mm in diam., abaxially with a pair of glands near base, 0.25–0.75 mm in diam. and touching the petiole or remote from petiole by up to 3 mm, additionally with 0–8 submarginal glands on each half of blade, 0.25–0.5 mm in diam. and 1–3 mm distant from margin. Inflorescences 1–8 cm long, sometimes bisexual but often only staminate flowers present, staminate part 8–10 mm in diam., very often reddish. Bracts of staminate cymules 0.4–0.75 mm long, with a pair of undivided glands 0.2–0.25(–0.5) mm long and distinctly overtopped by the bract. Staminate flowers 1 (rarely 3: NGF 28274) per cymule, pedicel (0.75–)1.5–2.5 mm long; sepals 2, c. 1 mm long; stamens (6–)8(–10) per flower, with filaments and anthers each c. 0.6 mm long. Pistillate flowers 1 per thyrse; pedicel 7–10 mm long; sepals 2, c. 0.75 mm long; ovary 2 mm long, bicarpellate, papillate, style often absent, 0–0.25(–1) mm long, stigma 3–6 by 0.3–0.6 mm, often widest at base and continuously narrowing, undivided, glandless or with small (c. 0.5 mm long) apical gland. Fruits 1 per infructescence; pedicel 1.5–3 cm long; calyx persistent; fruit excl. style 6–7 by 6–7 mm, papillate, sulcate, not carinate, style persistent; opened fruits not found, pericarp c. 0.25 mm thick (i.e. 1:25 of fruit length). Seeds often only one per fruit, c. 5.5 by 4.5 mm.
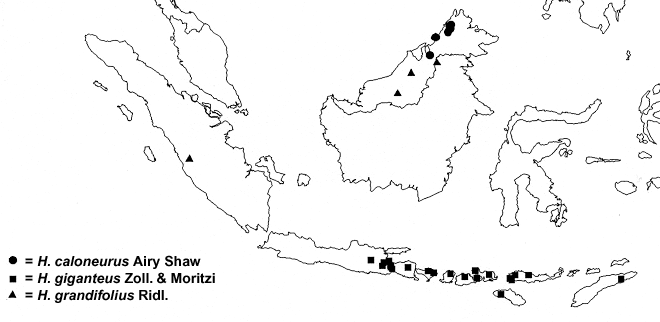
Distribution — Malesia: Moluccas (Morotai), New Guinea.
![]()
Habitat & Ecology — Found in mixed montane to subalpine forest, Nothofagus forest, moss forest, often along tracks, in regrowth and on forest margins, in scrub, grassland, or fire vegetation. Locally rare to common. Soil: grey clay, sand, peat, brownish soil, limestone. Altitude: 1,800–3,500 m (but compare H. minutiflorus). Flowering: January, February, May to October; fruiting all the year round.
Vernacular names — New Guinea: amp (Mendi); ambubori, ambuwaji (Enga); embo (Tari); kurup, kurupu (Hagen, Tomba); gugau (Chimbu, Masul).
Notes — 1. The exact date of publication of Nova Guinea 12 is not known. Therefore, it remains uncertain whether H. arfakiensis, published in October 1917, is indeed the oldest name. I follow Airy Shaw (1980a) in maintaining this name.
2. H. minutiflorus is united here with H. arfakiensis. Airy Shaw (1968) had admitted the close resemblance of both taxa, although he later (1980b) changed his opinion. H. minutiflorus is characterized by few quantitative characters, in particular smaller staminate flowers (e.g., only 4 stamens per flowers with anthers c. 0.25 mm long and filaments c. 0.2 mm long) and pistillate flowers with large stigmatic glands covering ± half of the stigma length. Comparable plants are also known from New Guinea, e.g. BW 14194, with floral bracts 0.4 mm long, pedicels of staminate flowers 0.75 mm long, a flower diameter of 1 mm, and only 6 stamens per flower. Additionally, type (Kostermans 884) and paratype (Kostermans 914) of H. minutiflorus were found at elevations of 100–250 m, very different from all specimens from New Guinea (see above).
Homalanthus caloneurus Airy Shaw, Kew Bull. 21 (1968) 413; Kew Bull. Add. Ser. 4 (1975) 135; Esser, Blumea 42 (1997) 433, Map 2. — Type: BRUN (Ashton) 2520 (K holo; iso L), Borneo, Brunei, Gunong Pagon Priok.
Homalanthus populneus (Geiseler) Pax var. cordifolius Heine, Feddes Repert. Spec. Nov. Regni Veg. 54 (1951) 235. — Type: J. & M.S. Clemens 32620 (M holo; iso B, BM, HBG, K-2 sheets, L, NY), Borneo, Sabah, Mount Kinabalu, Marai Parai.
Tree up to 20 m tall. Glabrous. Bark grey. Stipules 2–6 cm long. Leaves: petiole 3–10 cm long, apically with a pair of disc-shaped glands 0.5–1.25 mm in diam. situated laterally or more often adaxially; lamina ovate, 5–10 by 4.5–12 cm, index 0.8–1.1, base emarginate to truncate, not peltate, apex acuminate to cuspidate, lower surface whitish with very distinct, completely visible venation, side veins in 12–14 pairs below the apex, angle of divergence 45–55°, joined towards the margin, tertiary veins percurrent, quarternary veins reticulate, truly adaxial glands absent (but compare petiolar glands), abaxially with (0–)2–11 glands on each side of midvein, 0.25–0.5 mm in diam. and close to (often only 5–10 mm distant from) midvein. Inflorescences 7–10 cm long, bisexual but often without female flowers, staminate part 5–6 mm in diam. Bracts of staminate cymules c. 1 by 0.75 mm, with a pair of undivided glands c. 1 mm long and only slightly to not overtopped by the bract. Staminate flowers 3 (rarely 1: Clemens 34463) per bract; pedicel up to 1.5–2 mm long; sepals 2, c. 1.25 mm long; stamens 8 per flower, with filaments shorter than anthers and anthers c. 0.25 mm long. Pistillate flowers 4–20, at base of staminate thyrse or, rarely, intermixed with staminate flowers; pedicel 3–4 mm long; sepals 2, c. 0.5 mm long; ovary 3 mm long, bicarpellate, papillate, style 0.5–1 mm long, stigma 6–7 mm long, undivided, with an apical to subapical gland of 0.75–2 mm, apically up to 0.5 mm of stigma glandless. Fruits up to 20 per infructescence; pedicel 5–7 mm long; calyx persistent; fruit excl. style 7–8 by 6–7 mm, hardly sulcate, carinate at back, papillate or not, outer layer shrinkling, style persistent; opened fruit not seen. Seeds not studied, according to Airy Shaw (1968) 5–6 by c. 4 mm, with small arillode.
Distribution — Borneo (Sabah, Brunei).
 (circles on Borneo)
(circles on Borneo)
Habitat & Ecology — Found in disturbed montane forests, on ridge sides. Soil lateritic. Altitude: 1,200–2,500 m. Flowering: March, May, July to September; fruiting: March to May.
Vernacular names — Borneo: belantas, tapang lalat, ludai.
Note — H. caloneurus is unique in the genus in terms of the venation which is completely visible and conspicuous below, and by the laminar glands quite close to the midrib. The position of the petiolar leaf glands is variabel, in contrast to species like H. fastuosus with which Airy Shaw had compared it.
Homalanthus fastuosus (Linden) Fern.-Vill. in Blanco, Fl. Filip. ed. 3, Nov. App. (1880) 196; Vidal, Sin. Gen. Pl. Leños. Filip. Atlas (1883) 39, t. 84H; Revis. Pl. Vasc. Filip. (1886) 247; Merr. & Merritt, Philipp. J. Sc., Bot. 5 (1910) 357; Pax & K.Hoffm. in Engl., Pflanzenr. IV, 147, v (1912) 45; Merr., Enum. Philipp. Flow. Pl. 2 (1923) 459; Burkill, Dict. Econ. Prod. Malay Pen. 1 (1935) 1182; Salvosa, Lex. Philipp. Trees (1963) 105; Hsieh, Fl. Taiwan 3 (1977) 487, pl. 697; Airy Shaw, Alph. Enum. Euph. Philipp. Is. (1983) 32; Esser, Blumea 42 (1997) 435, Map 3. — Mappa fastuosa Linden, Belg. Hort. 15 (1865) 100; Cat. Pl. Exot. 19 (1865) 4. — Carumbium fastuosum (Linden) Müll.Arg. in DC., Prodr. 15.2 (1866) 1144; Baillon, Adansonia 6 (1867) 348, pl. 8. — Neotype (Esser, 1997): Elmer 17878 (L holo; iso BM, C, F, HBG, NY, P, U), Philippines, Luzon, Prov. Laguna, Los Baños (Mt. Maquiling).
Dibrachion peltatum Regel, Index Sem. Hortus Bot. Petrop. 1865 (1866) 51; Gartenflora 15 (1866) 100, t. 504. — Lectotype (Esser, 1997): Gartenflora 15 (1866) t. 504.
Homalanthus alpinus Elmer, Leafl. Philipp. Bot. 1 (1908) 307; Merr. & Merritt, Philipp. J. Sc., Bot. 5 (1910) 357; Pax & K.Hoffm. in Engl., Pflanzenr. IV, 147, v (1912) 48; Merr., Philipp. J. Sc., Bot. 9 (1915) 486; Enum. Philipp. Flow. Pl. 2 (1923) 459; Salvosa, Lex. Philipp. Trees (1963) 105; Airy Shaw, Kew Bull. 21 (1968) 414; Alph. Enum. Euph. Philipp. Is. (1983) 32. — Type: Elmer 7523 (holo: PNH, †; iso E, K, L, NY, WRSL), Philippines, Luzon, Tayabas Province, Lucban.
Homalanthus bicolor Merr., Philipp. J. Sc., Bot. 4 (1909) 282; Enum. Philipp. Flow. Pl. 2 (1923) 459; Salvosa, Lex. Philipp. Trees (1963) 105; Airy Shaw, Alph. Enum. Euph. Philipp. Is. (1983) 32. — Type: FB (Mearns & Hutchinson) 4722 (holo: PNH, †, fragm. A, photo A; iso K-2 sheets, NY), Philippines, Mindanao, Misamis Province, Mount Malindang.
Homalanthus milvus Airy Shaw, Kew Bull. 36 (1981) 611. — Type: BS (Ramos & Edaño) 26406 (K holo; iso A, P), Philippines, Luzon, Province Nueva Ecija, Mt. Umingan.
Tree up to 8(–18) m tall, dbh up to 30 cm, widely spreading and crookedly branched, with nearly flat-topped crown. Glabrous. Bark brown to grey-brown, marked with scattered corky pustules and long low horizontal ridges; outer bark soft, very thin, not detaching; inner bark soft, pale greenish-straw darkening to pale brownish on exposure. Stipules 1.5–8.5 cm long. Leaves densely crowded towards end of twigs; petiole 3–30 cm long, apically with a pair of abaxial-lateral, disc-shaped glands 0.6–2 mm in diam., rarely glandless or with another subapical pair of glands; lamina orbiculate to ovate, 4–23 by 3.5–24 cm, index 0.8–1.3, base cuneate to rounded to emarginate, peltate by up to 40 mm or not peltate, apex mucronate to acuminate, lower surface whitish between the veins, with most of the venation of different colour, side veins in 8–15 pairs below the apex, angle of divergence 45–60°, joined towards the margin, tertiary veins percurrent, quarternary veins reticulate to indistinctly percurrent, adaxially glandless, abaxially with 0–17 laminar glands on each half of blade, 0.25–0.75 mm in diam. and 5–30 mm distant from margin. Inflorescences 6–30 cm long, usually bisexual, staminate part 7–11 mm in diam. Bracts of staminate cymules c. 0.6–1 mm long, totally hidden by large pair of undivided glands 1–2 mm long. Staminate flowers 3–8 per cymule, often less than half of cymules 3-flowered; pedicel 1.5–3 mm long; sepals 2, c. 1 mm long; stamens 6–8 per flower, with filaments and anthers each c. 0.4–0.5 mm long. Pistillate flowers 6–45 per inflorescence; pedicel 3–5 mm long; sepals 2, c. 1 mm long; ovary 2 mm long, papillate, style 0.5–2 mm long, stigma 3–6 by 0.6–1 mm, undivided (rarely apically slightly divided: BS 13272), with quite small (0.75–2.5 mm long) apical gland. Fruits 6–30 per infructescence; bract often persistent; pedicel 4–13 mm long; calyx persistent, c. 1 mm long; fruit excl. style 6–9 by 5.5–8 mm, distinctly papillate, not sulcate, carinate and often with characteristical lateral, shallowly-conical projections and obovoid in shape, style persistent; regularly opened fruits not seen, pericarp c. 0.3–0.4 mm thick (£ 1:20 of fruit length). Seeds c. 3–5 by 3–4 mm, for a large part (at least the upper half) covered by arillode.
Distribution — Taiwan (Orchid Island, Botel Tobago) and Malesia: Philippines (Luzon to Mindoro and Panay, Negros, NW Mindanao).
![]()
Habitat & Ecology — Found on slopes and ridges, in hill forest, along trails or at exposed edges of dipterocarp forest, mossy forest, mountain forest, in disturbed areas of old garden sites, in rocky ravines, creekbanks, on rim of craters. Soil: sandy-loamy, limestone. Locally common, mostly above 750 m. Altitude: 50–2,600 m. Flowering: January to March, July to November; fruiting: all year round.
Uses — Used as fuel and firewood. The green smoke is said to make the eyes run unusually. The species is also known as fish poison (Burkill, 1935).
Vernacular names — Philippines: balanti (P. Vis., Tag.); botinag (Salvosa, 1963); buta (Tagalog; Salvosa, 1963); butabuta, butabutan abibilog (Ifugao); minunga; topi (Sub.; Salvosa, 1963).
Notes — 1. H. fastuosus was described from cultivated plants introduced into Botanical Gardens in Central Europe in the midth of the 19th century. A type collection has never been cited; even the exact locality of origin is not known. No possible type or surviving cultivation is present at BR and LG, where collections of Linden could be expected. Therefore, a neotype had to be chosen. I decided for the flowering collection with the largest number of available duplicates, which is Elmer 17878.
2. H. alpinus differs from H. fastuosus only by non-peltate leaves. This character is of limited value, like in other species of the genus. This may be shown by FB (Mearns & Hutchinson) 4722. The leaves of this collection are not or slightly (by up to 5 mm) peltate on the same twig. H. bicolor is based on this collection, separated from H. fastuosus primarily through the only slightly peltate laves. Pax & Hoffmann (1912) cited this specimen under H. fastuosus, a species they keyed out and characterized by peltate leaves. Other collections on which non-peltate and peltate leaves were found together are SF (Sinclair & Edaño) 9664 and Jacobs 7180. Collections with peltate and non-peltate leaves usually were gathered in the same localities, except few cases mentioned in note 4.
3. H. milvus was based on the unusual rhomboid leaf-shape with cuneate base. But this is only an extreme within the natural variation, and comparable leaf-shapes, although very rare, may be found in, e.g., H. populneus [BS (Ramos) 14460] or in Triadica sebifera (L.) Small [formerly Sapium sebiferum (L.) Roxb.], a species of another genus in the same tribe very similar in habit to several Homalanthus species.
4. Within the range of H. fastuosus some variation may be found, especially in leaf characters. In Taiwan, all collections studied may be recognized by very shortly acuminate, distinctly peltate leaves of remarkable similar size (6–8 by 6–8 cm). In the southernmost part of the range, e.g. in Mindanao, only slightly to not peltate leaves were found.
Homalanthus giganteus Zoll. & Moritzi, Natuur-Geneesk. Arch. Ned.-Indië 2 (1845) 584; Hassk., Flora 30 (1847) 662; Baillon, Étude Euphorb. (1858) 538, Atlas pl. 8, fig. 29–31; J.J.Sm. in Koord. & Valeton, Bijdr. Boomsoort. Java 12 (1910) 625; Pax & K.Hoffm. in Engl., Pflanzenr. IV, 147, v (1912) 51; Koord., Exkurs.-Fl. Java 2 (1912) 507; Backer & Bakh.f., Fl. Java 1 (1963) 498; Van Steenis, Mountain Fl. Java (1972) fig. 23, pl. 19-5; Airy Shaw, Kew Bull. 37 (1982) 24; Esser, Blumea 42 (1997) 438, Map 2. — Carumbium giganteum (Zoll. & Moritzi) Miq., Fl. Ind. Batav. 1.2 (1859) 687; Müll.Arg. in DC., Prodr. 15.2 (1866) 1145. — Type: Zollinger 2524 (A, BM, P), Java, prov. Malang, prope Gebok Klakka.
Homalanthus niveus Pax & K.Hoffm. in Engl., Pflanzenr. IV, 147, v (1912) 51; Airy Shaw, Kew Bull. 37 (1982) 25. — Type: Zollinger 1467 (holo? B, †), Lesser Sunda Islands, Bali, Tjator.
Tree up to 30 m tall, rapidly growing, dbh up to 45 cm. Twigs villose to glabrous. Bark grey to pale brown, finely cracked, can be rubbed off, living part 0.1–0.5 cm thick, inside yellow. Stipules 1–8 cm long. Leaves: petiole 3–34 cm long, glabrous at base, yellowish brown villose to glabrous at apex, at apex with 1–6 pairs of glands, 0.5–1.5 mm in diam., situated abaxially, laterally, or adaxially on petiole, or surrounding the petiole apex; lamina orbiculate to ovate, 3.5–34 by 4–28 cm, large in seedlings, smaller in older trees, index 0.75–1.2, base truncate to emarginate and often attenuate at the very base, peltate by 0.1–5 cm or not at all, apex mucronate to acuminate, upper surface glabrous, lower surface usually whitish, sometimes including the smallest veins, sometimes complete venation or only larger veins of different colour, rarely not whitish at all, yellowish brown villose to glabrous at base, especially above veins, otherwise glabrous, side veins in 10–14 pairs between petiole and the apex, angle of divergence 45–55°, joined towards the margin, tertiary veins percurrent, quarternary veins reticulate and often quite indistinct, adaxially glandless, abaxially with 0–5 laminar glands, c. 0.25 mm in diam. and close to the margin (e.g., 6 mm distant from margin). Inflorescences 8–15 cm long, bisexual, glabrous, staminate part 2–4 mm in diam. Bracts of staminate cymules 1–1.5 mm long, basally with a comparatively small pair of undivided glands 0.25–0.75 mm long and distinctly overtopped by the bract. Staminate flowers 1 per cymule; pedicel 1.25–1.5 mm long; calyx consisting of 1, conspicuously inclinate sepal c. 1 mm long; stamens (6–)10–15 per flower, with filaments and anthers each c. 0.5 mm long. Pistillate flowers 2–14 per thyrse; pedicel 8–22 mm long, shortest in distal flowers; sepals 2, c. 1.5 mm long; ovary 3 mm long, bicarpellate, papillate, style 1 mm long, stigma 3–4 by c. 0.5 mm, apically divided over at least half of its length, sometimes even over its whole length (and then apparently four per fruit), very rarely undivided, with apical gland c. 2 mm long. Fruits 2–14 per infructescence; bract persistent; pedicel 15–35 mm long; calyx persistent; fruit excl. style 7–11 by 8–9 mm, clavate in shape with 1–3 mm long, narrow base, papillate, hardly sulcate, each carpel distinctly carinate; style 2–4 mm long, stigma 3–5 mm long; shed unopened, but opening later loculicidally, pericarp c. 0.3 mm thick (i.e. less than 1:20 of fruit length), remaining columella c. 2 mm long, alate. Seeds c. 4.5 by 4 mm, upper half surrounded by arillode.
Distribution — Malesia: E Java, Lesser Sunda Islands.
(squares on Java and Lesser Sunda Islands)
Habitat & Ecology — Found as pioneer species in Casuarina forest, Podocarpus-Eugenia forest, montane forests, thickets, grassy areas, along ravines, on riverbanks. Locally very common. Soil: sand, loam, marl. Altitude: 350–2,400 m. Flowering: all year round; fruiting: May, August to October. Leaves deciduous (Smith, 1910).
Vernacular names — Lesser Sunda Islands, Bali: glantih. Flores: haju lente. Java (fide Smith, 1910): djarak prati gede, karembi, tundjung, tutup; usually the same names as known for H. populneus.
Uses — Dye for cloth.
Notes — 1. No type of Homalanthus niveus could be traced in any of the herbaria consulted for this study. Pax & Hoffmann (1912), however, cited no reliable diagnostic characters for their H. niveus. Pubescence and whitish colour of the lower leaf surface are indeed variable in H. giganteus, and the bifurcate stigmata are diagnostical for H. giganteus, whereas simple ones (as cited by Pax & Hoffmann) are exceptional but not unexpected (compare H. populneus). In most other characters, their descriptions of both species are identical. The synonymy of both taxa had already been suspected (with doubt) by Airy Shaw (1982), and he certainly was right.
2. In several characters, e.g., in indumentum, leaf base, measurements, and glands, H. giganteus is the most variable species of the genus. Leaves of seedlings may be peltate (Van Steenis 7305) or non-peltate (Schmutz 4080), and differences between young and old leaves have been noted by several authors, e.g., Van Steenis (1972). Additionally, there are some differences between plants of Java and Bali to those of the remaining Sunda Islands. In the former, the petiolar leaf glands are undivided (i.e., only one pair), and the fruits are somewhat larger (9–11 against 7–8 mm length).
3. Inflorescence buds are found quite often in H. giganteus, contrary to all other Malesian taxa. They are stiff, with densely packed bracts. This may be ecologically correlated with the deciduous leaves, also not known from any other Malesian species.
Homalanthus grandifolius Ridl., J. Fed. Malay States Mus. 8 (1917) 84; Esser, Blumea 42 (1997) 439, Fig. 1, Map 2. — Type: Robinson & Kloss s.n. (BM holo; iso BM, K), Sumatra, Gunung Koerintji.
![]()
![]()
![]()
Small shrub to medium tree. Glabrous. Stipules 2.5–7.5 cm long. Leaves: petiole 4.5–15 cm long, glandless; lamina orbiculate to ovate, 9.5–17 by 9.5–15.5 cm, index 1–1.3, base truncate to emarginate, distinctly peltate by 5–15 mm (not peltate: Burtt 12902), apex mucronate to acuminate, lower surface whitish with larger veins of different colour, side veins in 11–12 pairs below the apex, angle of divergence 50–55°, joined towards the margin, tertiary veins percurrent, quarternary veins percurrent to reticulate and quite indistinct, adaxially glandless, abaxially with a pair of large laminar glands 1–2 mm in diam. very close to and nearly touching petiole, additional laminar glands 4–26 on each half of blade, 0.3–1.5 mm in diam., in one or two rows or irregularly dispersed, nearly equally distant from margin and midrib or somewhat closer to margin. Inflorescences 9–14 cm long, bisexual, staminate part c. 7 mm in diam. Bracts of staminate cymules 1–1.25 mm long, nearly completely covered by a pair of undivided glands 1–1.5 mm long. Staminate flowers 1–3 per cymule; pedicel c. 2 mm long; sepals 2, c. 1 mm long; stamens 8–10 per flower, with filaments c. 0.3 mm long and anthers c. 0.5 mm long. Pistillate flowers 15–24 per thyrse; pedicel 4–8 mm long; sepals 2, 1–2 mm long; ovary 2 mm long, papillate, style 0.1–0.2 mm long, stigma 2.5–6 mm long, undivided, with apical to subapical gland 1–2 mm long. Fruits 11–15 per infructescence; pedicel 15–20 mm long; calyx caducous; fruit excl. style 7–8 mm long, papillate, style persistent, c. 0.75 mm long. Seeds not studied.
Distribution — Malesia: Sumatra, Borneo (Sabah, Sarawak, NE Kalimantan).
(triangles on Sumatra and Borneo)
Habitat & Ecology — In evergreen hill forest and in riverside vegetation, in gaps of fallen trees. Altitude: 1,400–2,400 m. Flowering and Fruiting: April, May, July.
Notes — 1. The holotype bears the number 212 written on the label, which may be the collection number.
2. The several Bünnemeijer collections from Sumatra are labelled 'Homalanthus sumatranus J.J.Sm.', a name which obviously has never been published.
3. This species has been omitted in Airy Shaw’s treatments of the Euphorbiaceae of Borneo and Sumatra (1975, 1981).
4. The collections from Borneo differ from the Sumatran plants in more elliptic leaves (index 1.2–1.3 instead of 1.0–1.1), shorter floral bracts totally hidden by their glands, and truly apical (instead of subapical) stigmatic glands. These differences do not justify taxonomic separation. The single collection with non-peltate leaves (Burtt 12902) agrees in all other characters with the remaining collections and is, therefore, attributed to this species.
Homalanthus longistylus Lauterb. & K.Schum. in K.Schum. & Lauterb., Fl. Schutzgeb. Südsee (1900) 407 [as 'longistilus']; Pax & K.Hoffm. in Engl., Pflanzenr. IV, 147, v (1912) 48; Airy Shaw, Kew Bull. Add. Ser. 8 (1980) 118; Esser, Blumea 42 (1997) 441, Map 4. — Neotype (Esser, 1997): Clemens 6603 (B holo; iso A), Papua New Guinea, Morobe Distr., Sattelberg, Yoangen, Mongi River.
Homalanthus papuanus Pax & K.Hoffm. in Engl., Pflanzenr. IV, 147, v (1912) 45; Airy Shaw, Kew Bull. 21 (1968) 409; Kew Bull. Add. Ser. 8 (1980) 117. — Type: Dr. Stephan s.n. (holo: B, †, fragm. and drawing WRSL), Papua New Guinea, Bismarck Archipelago, New Ireland, Muliama.
Carumbium populifolium auct. non Reinw.: Warb., Bot. Jahrb. Syst. 13 (1891) 352.
Carumbium populneum auct. non (Geiseler) Müll.Arg.: K.Schum., Notizbl. Königl. Bot. Gart. Berlin 2 (1898) 129.
Homalanthus populneus auct. non (Geiseler) Pax: K.Schum. & Lauterb., Fl. Schutzgeb. Südsee (1900) 407, p.p. quoad Lauterbach 1423; Airy Shaw, Kew Bull. 21 (1968) 409; Peekel, Fl. Bismarck Archip. for natural. (1984) 316, fig. 511.
Homalanthus novoguineënsis auct. non (Warb.) K.Schum.: Pax & K.Hoffm. in Engl., Pflanzenr. IV, 147, v (1912) 49, p.p. quoad Peekel 163, Schlechter 13761.
Homalanthus fastuosus auct. non (Linden) Fern.-Vill.: Airy Shaw, Kew Bull. 21 (1968) 409.
Homalanthus trivalvis auct. non Airy Shaw: Airy Shaw, Kew Bull Add. Ser. 8 (1980) 120.
Tree up to 12 m tall, dbh 20 cm. Glabrous (rarely youngest parts with yellowish-pale, c. 0.5 mm long hairs: NGF 12576). Bark reddish. Stipules 1.5–3 cm long. Leaves: petiole 8–12 cm long, apically with a pair of abaxial-lateral, disc-shaped glands 0.75–1.5 mm in diam.; lamina orbiculate to ovate, 5–17 by 4–16 cm, index 0.9–1.2, base rounded to slightly emarginate, not to distinctly (up to 10 mm) peltate, apex mucronate to acuminate, lower surface whitish with smaller veins of different colour, side veins in 10–13 pairs below the apex, angle of divergence 45–55°, joined towards the margin, tertiary veins percurrent, quarternary veins percurrent to slightly reticulate, adaxially glandless or with a pair of glands or one gland on lamina base c. 0.75 mm in diam., abaxially with 0–1(–14) glands on each side of midvein, 0.3–0.6 mm in diam. and 2–15 mm distant from margin with basal ones at least 10–15 mm distant from midvein. Inflorescences 7–12 cm long, often without pistillate flowers, but sometimes bisexual, staminate part 7 mm in diam. Bracts of staminate cymules 1–1.5 mm long, with a pair of undivided glands c. 0.5–1.25 mm long, always distinctly overtopped by the bract. Staminate flowers (1–)3 per cymule; pedicel 1.5–2 mm long; sepals 2, c. 0.75–1.25 mm long; stamens 6–8(–10) per flower, with filaments and anthers each c. 0.4 mm long. Pistillate flowers (4–)6–14 per inflorescence; pedicel 2 mm long; sepals 2, 0.5–0.75 mm long; ovary papillate, style nearly absent, stigma c. 2 by 0.4 mm, undivided, with apical gland c. 0.75 mm long. Fruits 4–7 per infructescence; bract persistent; pedicel 9–15 mm long; calyx persistent; fruit excl. style 6–8 by 8–9 mm or 12–14 by 12–15 mm (see note 3), papillate, sulcate, hardly to distinctly carinate, style persistent, 0.5–3 mm long, stigma 4–9 mm long; opened fruits not seen, pericarp c. 1 mm thick (slightly less than 1:10 of fruit length). Seeds c. 7 by 4 mm, upper half covered by arillode.
Distribution — Malesia: Papua New Guinea (in particular Sattelberg region and the Bismarck Archipelago).
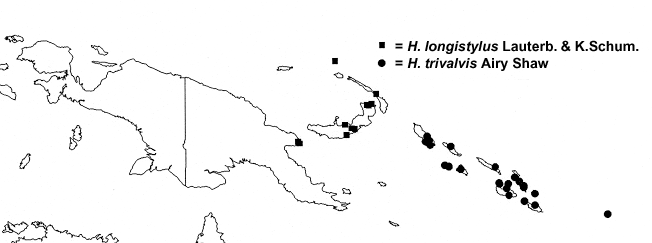 (squares on New Guinea and Bismarck Archipelago).
(squares on New Guinea and Bismarck Archipelago).
Habitat & Ecology — Found in secondary forests, open bush, banana and taro gardens, often as a major weed, and Nothofagus dominated moss forest. Soil: limestone. Locally uncommon to common. Altitude: 50–1,800 m. Flowering: January, March, July; fruiting: January, March, August, September.
Uses — The wood is used for building houses. The leaves are used for wrapping taro for cooking. The latex is irritant to the eyes. The terminal buds are eaten by women to induce abortion (Peekel, 1984).
Vernacular names — Papua New Guinea: a posoba (Namatanai, New Ireland); demeri (Sattelberg); kurip, pasok-mataves (Lamekot; Peekel, 1984); la pokakasi (W Nakani, New Britain); paba (Gazelle Peninsula); pasoba (Pala), pesem (Ugana); posom (Muliama); puba (Kuanua).
Notes — 1. The type of H. longistylus, Bamler II 1b, could not be traced in any of the herbaria consulted, including the Lauterbach herbarium (now WRSL). Therefore, a neotype is proposed here. Clemens 6603 was chosen because it agrees with the description of H. longistylus and is the only collection from the Sattelberg region (the type locality) that was found in more than one herbarium.
2. The type of H. papuanus is remarkable only by peltate leaves and 1-flowered staminate cymules. Both characters are found to be variable in such a way in H. longistylus that H. papuanus could easily be put into its synonymy.
3. The species is circumscribed here in particular by the abaxial-petiolar, disc-shaped leaf glands, bicarpellate ovaries, and bract glands small compared to the bracts itself. Pax & Hoffmann (1912) and Airy Shaw (1980a) cited the large, carinate fruits as characteristic for H. longistylus. This is not corroborated by the original literature. Schumann & Lauterbach (1900), when establishing the species from a single specimen, described no fruits, and obviously no later author studied the type, Bamler II 1b, not even Pax & Hoffmann. The large fruits described by them, repeated by Airy Shaw (1980a), probably were found in Lauterbach 1423 (WRSL), the only fruiting specimen they cited; this specimen had also been cited by Schumann & Lauterbach (1900) on the same page, but, instead, named H. populneus. The collections united now under H. longistylus obviously include two fruit types, but are indistinguishable otherwise. In most plants the fruits are small (6–8 by 8–9 mm) and hardly carinate. Only three collections bear large (12–14 by 12–15 mm), sharply carinate fruits, namely Lauterbach 1423 (Sattelberg), Clemens 41610 (Morobe Prov., Boana) and NGF (Croft & Katik) 41486 (New Britain). Because also H. papuanus is based only on a flowering plant, the taxonomic identity of the plants with large fruits remains uncertain; they are included here, with some doubt, under H. longistylus.
4. Without flowers, H. longistylus cannot not be distinguished from H. fastuosus. Therefore, the only collection from the Caroline Islands, Ledermann 13797 (B), a sterile one, remains undetermined.
5. Homalanthus trivalvis, again, is very similar to H. longistylus and distinguishable, besides characters of the staminate flowers, in particular by constantly tricarpellate ovaries and fruits. In H. longistylus, rarely single fruits may be tricarpellate, but never all on a twig, as noted by Peekel (1984). The citation of H. trivalvis for New Guinea by Airy Shaw (1980a) is erroneously based on such a specimen [NGF (Coode) 40469] of H. longistylus.
Homalanthus macradenius Pax & K.Hoffm. in Engl., Pflanzenr. IV, 147, v (1912) 51; Merr., Philipp. J. Sc., Bot. 9 (1915) 487; Enum. Philipp. Flow. Pl. 2 (1923) 459; Salvosa, Lex. Philipp. Trees (1963) 105; Airy Shaw, Alph. Enum. Euph. Philipp. Is. (1983) 33; Esser, Blumea 42 (1997) 443, Map 5. — Type: Elmer 10653 (holo: B, †; iso BM, E, F, HBG, K, L, NY, WRSL), Philippines, Mindanao, Davao, Todaya, Mt. Apo.
Homalanthus megaphyllus Merr., Philipp. J. Sc., Bot. 9 (1915) 485 (as 'Homalanthis'); Pax & K.Hoffm. in Engl., Pflanzenr. IV, 147, xiv, add. VI (1919) 56; Merr., Enum. Philipp. Flow. Pl. 2 (1923) 460; Salvosa, Lex. Philipp. Trees (1963) 105; Airy Shaw, Alph. Enum. Euph. Philipp. Is. (1983) 33. — Type: BS (Fénix) 15765 (holo: PNH, †, fragm. A, photo A), Philippines, Mindanao, Bukidnon Subprovince, Sumilao.
Homalanthus rotundifolius Merr., Philipp. J. Sc., Bot. 9 (1915) 486; Pax & K.Hoffm. in Engl., Pflanzenr. IV, 147, xiv, add. VI (1919) 57; Merr., Enum. Philipp. Flow. Pl. 2 (1923) 460; Salvosa, Lex. Philipp. Trees (1963) 106; Airy Shaw, Alph. Enum. Euph. Philipp. Is. (1983) 33. — Type: Ramos 1663 (holo: PNH,†; iso BM, L, NY, P), Philippines, Samar, Paranas.
Homalanthus surigaoënsis Elmer, Leafl. Philipp. Bot. 7 (1915) 2645; Pax & K.Hoffm. in Engl., Pflanzenr. IV, 147, xiv, add. VI (1919) 57. — Type: Elmer 14045 (holo: PNH, †; iso BM, C, E, F, HBG, K, L, NY, P, U), Philippines, Mindanao, Agusan Province, Cabadbaran (Mt. Urdaneta).
Homalanthus concolor Merr., Philipp. J. Sc. 20 (1922) 398; Enum. Philipp. Flow. Pl. 2 (1923) 459; Salvosa, Lex. Philipp. Trees (1963) 105; Airy Shaw, Alph. Enum. Euph. Philipp. Is. (1983) 32. — Type: BS (Ramos & Edaño) 39166 (holo: PNH, †; iso A, K), Philippines, Mindanao, Bukidnon Subprovince, near Tankulan.
Crooked, often shrub-like tree up to 15 m tall, dbh up to 15 cm, crown open and spreading. Glabrous. Bark pale grey-brown. Wood nearly odor- and tasteless. Stipules 1.5–12 cm long. Leaves: petiole 3–20 cm long, always with a pair of abaxial-lateral glands either cup-shaped, 0.5 mm in diam., or, more often, enlarged to peculiar foliaceous appendages, up to 6 mm long, with glandular margin and often irregularly divided; lamina ovate to orbiculate, 4–36 by 3.5–30 cm, index 1.0–1.2, base rounded and distinctly peltate by 5–120 mm, only rarely single leaves indistinctly peltate, apex rounded to acute to mucronate, lower surface often whitish with larger veins of different colour, but especially large leaves sometimes not whitish at all, side veins in 9–13 pairs between petiole and apex, angle of divergence 45–55°, only indistinctly joined towards the margin, tertiary veins percurrent, quarternary veins percurrent and often quite indistinct, adaxially glandless, abaxially without laminar glands. Inflorescences 10–21 cm long, bisexual, sometimes without female flowers, staminate part 5–14 mm in diam. Bracts of staminate cymules 0.3–1 mm long, completely covered by a pair of undivided glands 1–1.5 mm long. Staminate flowers 1 per cymule; pedicel 2–5 mm long; sepal 1, inclinate, 1.5–2 mm long; stamens c. 20–30 per flower, with filaments c. 0.5 mm long and anthers c. 0.3 mm long. Pistillate flowers 5–11 per thyrse; pedicel 2–5 mm long; sepal 1, inclinate, early caducous; ovary 2 mm long, bicarpellate, papillate, style 0.5–2.5 mm long, soon caducous and aborted near base, stigma 3–4 mm long, undivided, with apical gland 2–3 mm long. Fruits 5–11 per infructescence; bracts caducous; pedicel 4–9 mm long; calyx caducous; fruit 3.5–4.5 by 3.5–6 mm, papillate, deeply sulcate, each carpel slightly carinate, always without style; opened fruits rarely seen, pericarp c. 0.15 mm thick (c. 1:25 of fruit length), remaining columella alate, c. 1 mm wide. Seeds c. 4 by 3.5 mm, with apical arillode.
Distribution — Malesia: Philippines (E and SE part only: Negros and Samar to Mindanao).
![]()
Habitat & Ecology — Found in secondary forests from early to advanced stages, in forest edges, open places, and along streams. Soil dry, red, stony. Locally common. Altitude: 45–1,800 m. Flowering and fruiting: March to October with maxima in March to May and September to October.
Uses — The leaves are used for wrapping and covering food.
Vernacular names — Philippines: balalanti (Tagalog); balanti; balanting-bilog, Mindanao balanti (Fil.: Salvosa, 1963); biante (Cebuano); gibulag, glabulag; labagti, labugti, labulti, liuti, maquing, salingbaga (Bukidnon).
Notes — 1. Contrary to most other species, the whitish colour of the lower leaf surfaces may be absent; this is especially true for the largest leaves studied (e.g., the type of H. megaphyllus), but, less often, also for smaller leaves (e.g., the type of H. concolor).
2. H. megaphyllus is only a gigantic form of H. macradenius. The remarkable variability of its leaf size has been noted by Elmer (1915, under H. surigaoënsis), but is not greater than in some other species, e.g., H. giganteus.
Homalanthus nervosus J.J.Sm. in Lorentz, Nova Guinea 8 (1912) 792, t. 141; Pax & K.Hoffm. in Engl., Pflanzenr. IV, 147, vii, add. V (1914) 420; Airy Shaw, Kew Bull. 21 (1968) 412; Kew Bull. Add. Ser. 8 (1980) 118; P. van Royen, Alpine Fl. New Guinea 3 (1982) 2182, fig. 663; Esser, Blumea 42 (1997) 445, Map 6. — Type: von Römer 1241 (BO holo, n.v.; iso L-2 sheets), New Guinea, Irian Jaya, Gipfel des Hellwig-Gebirges.
Homalanthus vernicosus Gage in Herderschee, Nova Guinea 12 (1917) 483, t. 185; Pax & K.Hoffm. in Engl., Pflanzenr. IV, 147, xvii, add. VII (1924) 199; Airy Shaw, Kew Bull. Add. Ser. 8 (1980) 120. — Lectotype (here proposed): Pulle 711 (BO holo n.v.; iso K, L, U), New Guinea, Irian Jaya, Mount Hellwig.
Homalanthus deltoideus Airy Shaw, Kew Bull. 34 (1980) 594; Kew Bull. Add. Ser. 8 (1980) 117. — Type: Brass 30586 (K holo; iso L), Papua New Guinea, Eastern Highlands District, Mount Wilhelm, E slopes.
Tree up to 10(–16) m tall, dbh 15 cm, heavily branched and spreading, with umbrella-shaped crown. Glabrous. Outer bark brown to grey, with numerous small, light-coloured lenticels and discontinuous fissures; middle bark light green; inner bark straw to creamy. Stipules 1.2–5 cm long. Leaves: petiole 1.5–13 cm long, glandless; lamina ovate, 4–17 by 3–14 cm, index 1.1–2.4, base rounded to slightly emarginate, very base often attenuate, not or indistinctly (by 1 mm) peltate, apex acuminate to cuspidate, lower surface usually shiny and not whitish (rarely whitish beneath: `H. deltoideus'), 9–13 pairs of side veins below the apex, angle of divergence 45–65°, indistinctly to distinctly joined towards the margin, tertiary veins percurrent, quarternary veins percurrent to reticulate, adaxially with an undivided, disc-shaped, prominent gland on junction lamina base/petiole, 1–2 mm in diam., abaxially with a pair of conspicuous glands on lamina base, 0.75–3 mm in diam. and (0–)2–11 mm distant from midvein and petiole apex, sometimes absent on single leaves, without additional laminar glands (4 additional glands, 0.4 mm in diam.: Webster & Hildreth 15117). Inflorescences 2–18 cm long, often only with staminate flowers, if bisexual often with pistillate part longer than staminate part, staminate part 4–6 mm in diam. Bracts of staminate cymules 0.75–1.5 mm long, with a pair of undivided and often conspicuously shiny glands 0.6–1 mm long, distinctly overtopped by the bract. Staminate flowers 3 (rarely up to 6: Jacobs 8609) per cymule; pedicel 0.75–2 mm long; sepals 2, c. 0.75 mm long; stamens (6–)8 per flower, with filaments c. 0.25-0.5 mm long and anthers c. 0.5 mm long. Pistillate flowers 3–22 per thyrse; pedicel 2–16 mm long, shorter in distal flowers; sepals 2, c. 0.5–1 mm long; ovary 1–1.5 mm long, bicarpellate, papillate or non-papillate, style 0.25–2.5 mm long, stigma 2–4 by c. 0.5 mm, undivided, with a small apical gland 1–2 mm long. Fruits 2–7 per infructescence; bract persistent; pedicel 8–25 mm long; calyx persistent; fruit excl. style 3.5–6 by 4–5.5 mm, indistinctly papillate, slightly sulcate, not carinate, style persistent, often only 1-seeded with lateral style; opened fruits not seen, usually shed unopened, pericarp c. 0.1 mm thick (i.e. 1:40 of fruit length); remaining columella c. 4 by 0.5 mm, hardly alate. Seeds c. 3 by 2.25 mm, with apical arillode.
Distribution — Malesia: throughout New Guinea.
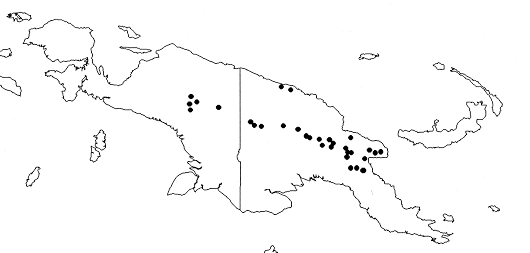
Habitat & Ecology — Found especially on ridges and slopes, in mossy, montane forest, Nothofagus forest, Castanopsis-oak forest, especially on margins, along tracks, in young (3–5 years old) or advanced (50 years old) regrowth, in open low scrub, on rocky streamsides. Soil: black peat over yellow clay, stony clay, loam, limestone, brown soil (pH 6–6.5), sand, of good to medium drainage or quite wet. Locally very common, sometimes even one of the principal species. Altitude: 1,400–2,600 m. Flowering and fruiting all year round, flowers with a maximum in July to August.
Uses — The fruits are favourite food for pigeons. The sap is known as fish poison and to be irritant to the eyes.
Vernacular names — New Guinea: amb (Mendi); ambobai, ambupi, ambupokai (Enga); bobo, gioriba (Middle Waria); bufan tikik (Telefomin); kareim (Aiyura); korosomore (Naho); kurup (Hagen, Wankl); mauk (Jal, Madang); nenifi (Wapi); tukko (Anona).
Notes — 1. H. nervosus and H. vernicosus are perfectly identical. Airy Shaw (1980a) distinguished both only by the leaf venation being conspicuously prominent above in the latter and not prominent in the former. It is correct that the type of H. vernicosus is untypical in this respect for the majority of the known specimens, but this is also true for the type of H. nervosus, a specimen not examined by Airy Shaw.
2. H. deltoideus differs from typical H. nervosus only in the leaves abaxially slightly whitish and in quite small inflorescences and flowers, e.g., only 6 stamens per flower. Also, the fruits are somewhat larger than in H. nervosus (8 by 7 mm). Airy Shaw himself (1968) had discussed Brass 30586, which he later made the type of H. deltoideus, concluding that it should not be separated from H. nervosus, not even by varietal name. This seems agreeable. Nevertheless, the most obvious character of H. nervosus, the leaves being shiny and not whitish beneath, shows exceptions.
Homalanthus novoguineënsis (Warb.) K.Schum. in K.Schum. & Lauterb., Fl. Schutzgeb. Südsee (1900) 407 [as 'novo-guineensis']; J.J.Sm. in Lorentz, Nova Guinea 8 (1912) 792; Pax & K.Hoffm. in Engl., Pflanzenr. IV, 147, v (1912) 48; Ridl., Trans. Linn. Soc., Bot. 9 (1916) 147; Airy Shaw, Kew Bull. 21 (1968) 410; Kew Bull. Add. Ser. 8 (1980) 118; Muelleria 4 (1980) 238, 244; Kew Bull. 35 (1981) 642, fig. 3B; Kew Bull. 37 (1982) 25; P.I.Forster, Telopea 6 (1994) 170; Esser, Blumea 42 (1997) 447, Map 7. — Carumbium novoguineënse Warb., Bot. Jahrb. 18 (1893) 199. — Type: Hellwig 555 (holo: B, †; iso K), Papua New Guinea, Finschhafen, am Sattelberg.
Homalanthus brachystachys Pax & K.Hoffm. in Engl., Pflanzenr. IV, 147, v (1912) 47; Airy Shaw, Kew Bull. 21 (1968) 410. — Lectotype (here proposed): Schlechter 16392 (A holo; iso P-3 sheets), Papua New Guinea, Kaiser Wilhemsland, am Minjem, 200 m.
Homalanthus tetrandrus J.J.Sm. in Lorentz, Nova Guinea 8 (1912) 791, t. 140; Pax & K.Hoffm. in Engl., Pflanzenr. IV, 147, vii, add. V (1914) 420; Airy Shaw, Kew Bull. Add. Ser. 8 (1980) 120; Kew Bull. 37 (1982) 25. — Type: von Römer 912 (BO holo, n.v.; iso L), New Guinea, Irian Jaya, Vorgebirge des Hellwig-Gebirges, 750 m.
Homalanthus elegans Gage in Herderschee, Nova Guinea 12 (1917) 483, t. 186; Pax & K.Hoffm. in Engl., Pflanzenr. IV, 147, xvii, add. VII (1924) 198; Airy Shaw, Kew Bull. 21 (1968) 410. — Type: Pulle 249 (BO holo, n.v.; iso K, L, U), New Guinea, Irian Jaya, North River near Kloofbivak, 30 m.
Homalanthus crinitus Gage in Herderschee, Nova Guinea 12 (1917) 484; Pax & K.Hoffm. in Engl., Pflanzenr. IV, 147, xvii, ad. VII (1924) 198; Airy Shaw, Kew Bull. Add. Ser. 8 (1980) 120. — Type: Pulle 168 (BO holo, n.v.; iso K, L), New Guinea, Irian Jaya, North River near Kloofbivak, 30 m.
Homalanthus beguinii J.J.Sm., Bull. Jard. Bot. Buitenz., ser. 3.6 (1924) 98; Heyne, Nutt. Pl. Ned. Ind. 1, 3rd ed. (1950) 959; Airy Shaw, Kew Bull. 37 (1982) 24. — Lectotype (here proposed): Beguin 1160 (BO n.v.; iso L), Moluccas, Ternate, Toramadiahi, 450 m.
Homalanthus pachystylus Airy Shaw, Kew Bull. 34 (1980) 595; Kew Bull. Add. Ser. 8 (1980) 119. — Type: Hyndman 4 (K holo; iso L), Papua New Guinea, Western District, Kiunga Subdistrict, 43 km NE Ningerum, 510 m.
Tree up to 25 m tall, dbh up to 50 cm, straight, much-branched and with umbrella-shaped crown; without buttresses. Glabrous to yellowish-brownish puberulous. Bark glabrous to puberulous, c. 5 mm thick, medium tan to creamy-white to grey, finely vertically fissured, with round pustular lenticels; under bark light green; inner bark straw to brown to purple-brown. Stipules 0.7–2 cm long. Leaves: petiole 2–13 cm long, glabrous to puberulous, glandless; lamina orbiculate to ovate to elliptic, 4.5–19 by 3.5–16 cm, index 0.8–1.7, base cuneate to rounded to slightly emarginate, peltate (up to 3 mm) or not, apex acute to acuminate, lower surface whitish but most of venation of different colour, glabrous to puberulous, side veins in 9–13 pairs below the apex, angle of divergence 45–55°, hardly joined towards the margin, tertiary veins percurrent, quarternary veins reticulate, adaxially on junction lamina base/petiole with an undivided, disc-shaped, prominent gland 1–2 mm in diam., glandless only when base peltate, abaxially usually with a pair of glands near base, 0.25–1.25 mm in diam. and 1–15 mm distant from petiole apex, additionally with 0–3 glands on each half of blade, 0.3–0.4 mm in diam. and 3–5 mm distant from leaf margin, rarely completely glandless abaxially. Inflorescences 5–13 cm long, sometimes bisexual and then pistillate part as long as or longer than staminate part, but quite often only with staminate or only with pistillate flowers, staminate part 5–7 mm in diam., glabrous to puberulous. Bracts of staminate cymules 0.9–1.25 mm long, at base with a cluster of foveolate to cup-shaped glands with shiny center and thickenend, glaucous-papillate margin, 0.75–1.5 mm long and consisting of at least 6 single glands 0.2–0.4 mm in diam., distinctly (by 0.4–0.8 mm) overtopped by the bract. Staminate flowers (1–)3(–6) per cymule; pedicel 1–2.5 mm long; sepals 2, c. 0.5–0.7 mm long; stamens 6–8 per flower, with filaments c. 0.3 mm long and anthers 0.3–0.4 mm long. Pistillate flowers 8–30 per thryse if not absent at all; pedicel 2–10 mm long; sepals 2, c. 1 mm long; ovary 1–2 mm long, bicarpellate, papillate, style 0.2–1 mm long, stigma c. 1–2 by 0.3–0.5 mm, undivided, with a minute apical gland (up to 0.5 mm long) or rarely glandless. Fruits 2–30 per infructescence; bract persistent; pedicel 0.8–3.5 cm long; calyx caducous; fruit excl. style 5–13 by 6–9 mm, papillate, distinctly to indistinctly sulcate, not to slightly carinate, style persistent, fruits often one-seeded and then with lateral, oblique style; regularly opened fruits not uncommon, pericarp 0.2–0.25 mm thick (i.e. 1:30 of fruit length), remaining columella 5 by 1 mm, slightly alate. Seeds c. 4.5 by 4–4.5 mm.
Distribution — N and NW Australia (West Australia, Northern Territory, Queensland) and in Malesia: Lesser Sunda Islands (eastern part), Moluccas, New Guinea, Solomon Islands.
![]()
Habitat & Ecology — Found in secondary growth of e.g. Castanopsis-oak forest, on plains and slopes, as landslide coloniser, in mossy forest with Myrtaceae, Papuacedrus, and Podocarpaceae, old garden sites, Eucalyptus savanna, ladang along shallow river, on riverbanks and by streams, forest edge along sago swamps, in low-lying and mostly water-logged fellows. Soil: old coral limestone, well-drained volcanic soil, dark marshy soil, rocky alluvium, poorly drained consolidated alluvium, clayey soil on limestone, alluvial sandy clay. Locally common, often even as characteristic and chief dominant species. Altitude: 10–2,300 m. Flowering and fruiting all year round, most often May to June. The fruit is said to be attractive to birds.
Uses — The latex is known as poison and harmful to the eyes. The sap of young leaves is administered to young children as mild laxative (Heyne, 1950). Leaves and bark are used as black dye (Heyne, 1950). The leaves are used for wrapping food, the wood as firewood, and for construction.
Vernacular names — Lesser Sunda Islands: ba’an (Dawan, Timor); bussi, fkeno (Timor); lalaa, tawa (Alor). Moluccas: palake, palate (Halmaheira). Irian Jaya: kauw (Maibrat, Vogelkop); kopa (Kapaukoe, Wissel Lakes). Papua New Guinea: ambuboi (Enga); as defen, defen (Miyanmin); bangiumba (Waskuk); bobo (Garaina, Bulolo); demeng sesakele (Finschhafen); hubawo (Kutubu); imbo (Huli); nenum (Wapi, Marok); simsam (Weng); ugafo (Orokaiva, Mumuni).
Notes — 1. For the distinction from H. populifolius see note 3 under that species.
2. H. pachystylus was distinguished from H. novoguineënsis by somewhat larger (7–13 mm long), non-carinate fruits with thicker styles. Airy Shaw admitted that there was no clear morphological gap between both taxa. Indeed, leaves and flowers of both taxa are identical, and the cited difference in the style could not be verified. Both taxa are united here. In this broad circumscription the fruits of H. novoguineënsis are more variable than those of the other Malesian species.
3. H. tetrandrus (including H. crinitus) differs from typical H. novoguineënsis only in the presence of a distinct indumentum, but is identical in all other characters studied. Usually, H. novoguineënsis is glabrous. Pubescence may be found, however, in most parts of its distributional area, including Australia. Density, length, and localization of the hairs is very variable. Hoogland 10417 and Schlechter 16392 show minute, 0.1 mm long and nearly papillate hairs only on the petiole apex; Brass 2006 and Lauterbach 2487 have somewhat longer hairs on petioles and midvein of leaves; the leaves of von Römer 912, the type of H. tetrandrus, are densely covered with c. 0.6 mm long hairs over the whole lower surface, in particular on the veins; most extreme is Hoover 494, where not only leaves but also stipules and floral bracts are densely hairy. It is suggested here to treat the hairiness as variable; consequently, the respective taxa have to be united.
Homalanthus populifolius Graham, Edinb. New Philos. J. 3 (1827) 175; Bot. Mag. 54 (1827) tab. 2780; Baillon, Étude Euphorb. (1858) Atlas pl. 8, fig. 22–26; Pax in Engl. & Prantl, Nat. Pflanzenfam. 3.5 (1890) fig. 60 (as ‘populneus’); Pax & K.Hoffm. in Engl., Pflanzenr. IV, 147, v (1912) 46, fig. 7; Airy Shaw, Kew Bull. 21 (1968) 410; Sykes, New Zeal. J. Bot. 7 (1969) 302; Airy Shaw, Kew Bull. Add. Ser. 8 (1980) 119; Muelleria 4 (1980) 238, 244; Kew Bull. 35 (1981) 642; P.I.Forster, Telopea 6 (1994) 169; Esser, Blumea 42 (1997) 450, Map 8. — Carumbium populifolium (Graham) Benth., Fl. Austral. 6 (1873) 150. — Type: Graham s.n. (E holo, photo in K; iso K, n.v.), native of Australia, from seed collected by Fraser in 1824, cultivated in Hort. Bot. Edinburgh and flowering in 1827.
Homalanthus goodenoviensis Airy Shaw, Kew Bull. 21 (1968) 417; Kew Bull. Add. Ser. 8 (1980) 118. — Type: Brass 24932 (K holo; iso L), Papua New Guinea, Milne Bay District, Goodenough Island (D'Entrecasteaux Group), 900 m.
Homalanthus populneus auct. non (Geiseler) Pax: Müll.Arg. in DC., Prodr. 15.2 (1866) 1144, pro parte (under Carumbium); De Wild., Icon. Horti Then. 5 (1904) 37, pl. 169.
Shrub or tree, up to 12 m tall, dbh 5 cm, with straight bole and open, spreading crown. Glabrous. Stipules c. 1.5 cm long. Leaves: petiole 1.5–7 cm long, glandless; lamina orbiculate to ovate, 3–11.5 by 2.5–9 cm, index 1.1–1.8, base obtuse, very base often attenuate, not peltate, apex acuminate, lower surface whitish with most of venation of different colour, side veins in 9–13 pairs below the apex, angle of divergence 45–60°, hardly joined towards the margin, tertiary veins percurrent, quarternary veins reticulate, adaxially with an undivided, disc-shaped, prominent gland on junction lamina base/petiole, 0.75–1 mm in diam., abaxially with 0–2 laminar glands c. 0.25 mm in diam. and c. 2–4 mm distant from leaf margin, basal ones not enlarged. Inflorescences 4–9 cm long, bisexual, staminate part c. 6 mm in diam. Bracts of staminate cymules 0.75–1 mm long, with a comparatively small pair of roundish to pillow-shaped glands c. 0.25(–0.6) mm long, distinctly overtopped by the bract, sometimes with 2–3 pairs of glands, rarely glandless (most bracts of Brass 28722). Staminate flowers 1(–3) per cymule; pedicel 0.75–2 mm long; sepals 2, 0.6–1 mm long; stamens 6–8 per flower, with filaments c. 0.3 mm long and anthers c. 0.4 mm long. Pistillate flowers 1–4 per thyrse; pedicel c. 5–15 mm long; sepals 2, 0.75–1.25 mm long; ovary 1.5–2 mm long, bicarpellate, papillate or glabrous, style c. 0.25–1 mm long, stigma 1–4 by 0.4–0.5 mm, undivided, with a minute apical gland c. 0.4 mm long or rarely glandless. Fruits 1–4 per infructescence; bract sometimes persistent; pedicel 1.2–3.8 cm long; calyx hardly persistent; fruit excl. style 5–8 by 6–8 mm, sulcate, not carinate, papillate or not, style persistent; regularly opened fruits not seen. Seeds not studied in Malesian specimens.
Distribution — NE and E Australia (Queensland, New South Wales), Solomon Islands and Malesia: New Guinea (Papua New Guinea, including Admirality Islands). Cultivated in many Botanical Gardens, and in Sri Lanka probably naturalized from cultivation.
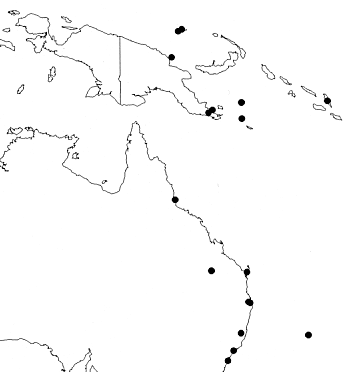
Habitat & Ecology — Found in lowland and lower montane forest regrowth, secondary forest, well drained or swamp forest, on ridges, riversides. Soil: coral rubble, sandy soil, brown loam. Locally occasional to common. Altitude: sea level up to 1,000 m. Flowering: July to January; fruiting: March, July, October to December. Flowers reported to have faint smell.
Uses — In Australia used as a pioneer tree in landscape rehabilitation projects. In Botanical Gardens planted as an ornamental tree.
Vernacular names — Papua New Guinea: kinapuk. Australia: native bleeding heart, native poplar, queensland poplar.
Notes — 1. Forster (1994) had united H. populifolius with H. nutans (G.Forst.) Guillemin from several of the Pacific Islands, the latter being the older name and taking priority. This is not followed here. H. nutans, instead, differs from the former in several significant characters, e.g., leaves with a pair of adaxial base glands, bract glands large and hardly overtopped by bract, staminate flowers with only 1 sepal and 10–20 stamens each, ovaries and fruits 2- or 3-carpellate. This is certainly enough to keep the two species separate. Pax & Hoffmann (1912) even put them in different sections of Homalanthus. The ovaries, however, may be papillate or not in both taxa and are, therefore, of minor significance (this character had been stressed by Forster). The superficial similarity in habit may also be found in several other species of the genus.
2. Homalanthus goodenoviensis, i.e. the collections from the Goodenough Island, differs from the remaining Malesian collections in the non-papillate ovaries and fruits and in the short pedicels of the staminate flowers. These pedicels are so short that the flowers remain covered partly by the bract; but, indeed, the flowers are not sessile as Airy Shaw (1980a) had stated. Airy Shaw (1968), when establishing this species, did not compare it with H. populifolius, and it is indeed not different from the latter.
3. Homalanthus populifolius is very close to H. novoguineënsis. Both are the only species of the genus where multiple bract glands may occur, and leaves, flowers, and fruits are hardly distinguishable. The stigma length, used repeatedly by Airy Shaw, does not separate both taxa. Interestingly, he first (1968) did not see a difference in this character, whereas he later (1981) distinguished H. populifolius by elongate stigmata. This character is as variable in H. populifolius as in most other species. Only two herbarium characters remain for separation from H. novoguineënsis, the number of bract glands (1–3 pairs against usually at least 6 pairs), and their morphology (roundish pillow-shaped with uniform surface against cup-shaped to foveolate with papillate margin and shiny center). Additionally, H. novoguineënsis sometimes is puberulous, contrary to H. populifolius, the number of pistillate flowers per thyrse is usually much higher in H. novoguineënsis, and inflorescences are more often unisexual in the latter; but these last characters are not always visible. The limits of both species remain somewhat vague, and field studies may be necessary to evaluate the reliability of these features. Airy Shaw (1968) suspected introgression. The hairy-papillate ovary surface, used by Forster (1994), does not distinguish both. Contrary to the repeated confusion with H. populneus (compare the literature citations under both taxa!) and remarks like those of Airy Shaw (1968), this species does not seem to resemble H. populifolius closely.
4. Malesian specimens differ from the 'typical' H. populifolius, like the forms found in Australia and in botanical gardens, in pistillate flowers with somewhat shorter stigmata (often only 1–2 mm long) and mostly only 1-flowered staminate cymules. Otherwise, they match the type well. The bract glands may be missing on individual bracts, but never on all. Somewhat exceptional is the collection NGF 36728; it is the only Malesian one with 3-flowered staminate cymules, the bract glands are 0.6 mm in diameter (the largest found in Malesia).
Homalanthus populneus (Geiseler) Pax in Engl. & Prantl, Nat. Pflanzenfam. 3.5 (1890) 96, excl. fig. 60; Kuntze, Revis. Gen. Pl. 2 (1891) 609; Merr., Philipp. J. Sc. 1 Suppl. (1906) 83; J.J.Sm. in Koord. & Valeton, Bijdr. Boomsoort. Java 12 (1910) 621; Pax & K.Hoffm. in Engl., Pflanzenr. IV, 147, v (1912) 46; Merr., J. Str. Br. Roy. As. Soc. (1921) 347; Enum. Philipp. Flow. Pl. 2 (1923) 460; Philipp. J. Sc. (1926) 387; Pl. Elmer. Born. (1929) 166; Corner, Ways. Trees Malaya 1 (1940) 257; Holthuis & Lam, Blumea 5 (1942) 202; Heyne, Nutt. Pl. Ned. Ind. 1, 3rd ed. (1950) 959; Heine, Feddes Repert. Spec. Nov. Regni Veg. 54 (1951) 235; Salvosa, Lex. Philipp. Trees (1963) 105; Backer & Bakh.f., Fl. Java 1 (1963) 498; Airy Shaw, Kew Bull. 26 (1972) 281; Whitmore, Tree Fl. Malaya 2 (1973) 102; Airy Shaw, Kew Bull. Add. Ser. 4 (1975) 136; Kew Bull. 36 (1981) 310; Kew Bull. 37 (1982) 25; Alph. Enum. Euph. Philipp. Is. (1983) 33; Keßler & Sidiyasa, Tropenbos Ser. 7 (1994) 129, fig. 96; Esser, Blumea 42 (1997) 453, Map 9; Esser & Welzen in Welzen & Chayam., Fl. Thailand 8, 2 (2007) 334, Fig. 7. — Stillingia populnea Geiseler, Croton. Monogr. (1807) 80. — Carumbium populneum (Geiseler) Müll.Arg. in DC., Prodr. 15.2 (1866) 1144. — Homalanthus populneus (Geiseler) Pax var. genuinus Pax & K.Hoffm. in Engl., Pflanzenr. IV, 147, v (1912) 46, nom. illeg. — Type: Herb. Thunberg in herb. Dahl s.n. (C holo, microfiche 2201-73-16 &-17; ?iso UPS, n.v.), s.loc. (see note 1).
Carumbium populifolium Reinw., Elench. Sem. Hort. Leiden ex Isis 1 (1823) 319 (nomen semi-nudum, see note 2); in Blume, Cat. Pl. Bogor. (1823) 105, nomen; T.Nees, Flora 8 (1825) 103, nomen (as `Garumbium'); Reinw., Syll. Pl. Nov. 2 (1825) 6; Miq., Fl. Ind. Batav. 1.2 (1859) 414; Scheff., Ann. Mus. Bot. Lugduno-Batavum 4 (1869) 127. — Type: Reinwardt s.n. (L holo?), Java.
Homalanthus leschenaultianus A.Juss., Euphorb. Gen. (1824) t. 16 fig. 53; Bl., Bijdr. 12 (1825) 627; Hassk., Cat. Hort. Bot. Bogor. (1844) 234. — Type: Leschenault de la Tour s.n. (P-JU Cat. 16568 holo, microfiche 1199/5), Java.
Excoecaria laevis Blanco, Fl. Filip. (1837) 788; Fl. Filip. ed. 2 (1845) 543; Fl. Filip. ed. 3 (1879) 194. — Homalanthus populneus (Geiseler) Pax var. laevis (Blanco) Merr., Sp. Blancoan. (1918) 230; Enum. Philipp. Flow. Pl. 2 (1923) 460 (`levis'); Salvosa, Lex. Philipp. Trees (1963) 106 (`levis'); Airy Shaw, Alph. Enum. Euph. Philipp. Is. (1983) 33. — Neotype (here proposed): Merrill Species Blancoanae 75 (L holo; iso BM, F, K, NY, P), Philippines, Luzon, Batangas Prov.
Carumbium populneum (Geiseler) Müll.Arg. var. minus Müll.Arg. in DC., Prodr. 15.2 (1866) 1145. — Homalanthus populneus (Geiseler) Pax var. minor (Müll.Arg.) Merr., Philipp. J. Sc., Bot. 7 (1912) 390; Pax & K.Hoffm. in Engl., Pflanzenr. IV, 147, vii, add. V (1914) 420; Merr., Sp. Blancoan. (1918) 230; Airy Shaw, Alph. Enum. Euph. Philipp. Is. (1983) 33. — Type: Meyen s.n. (holo: B, †), Philippines, Luzon, prope Manila.
Homalanthus populneus (Geiseler) Pax var. siccus Pax in Engl., Pflanzenr. IV, 147, v (1912) 46, p.p. quoad synonyma Excoecaria laevis, Carumbium populifolium var. minus; Merr., Philipp. J. Sc., Bot. 7 (1912) 390.
Homalanthus sulawesianus Airy Shaw, Kew Bull. 35 (1980) 398; Kew Bull. 37 (1982) 25. — Type: Musser 917 (K holo; iso K), Central Celebes, Sungai Sadaunta.
Homalanthus populifolius auct. non Graham: Fern.-Vill. in Blanco, Fl. Filip. ed.3, Nov. App. (1880) 196; Vidal, Revis. Pl. Vasc. Filip. (1886) 247; Hook.f., Fl. Brit. India 5 (1888) 469; Ridl., J. Fed. Malay States Mus. 8 (1917) 84; Fl. Malay Penins. 3 (1924) 313; Burkill, Dict. Econ. Prod. Malay Pen. 1 (1935) 1182.
? Homalanthus giganteus auct. non Zoll. & Moritzi: Boerl., Fl. Ned. Ind. 3 (1900) 295; Merr., J. Str. Br. Roy. As. Soc. (1921) 347.
Sapium sebiferum auct. non (L.) Roxb.: Hutch., J. Linn. Soc., Bot. 42 (1878) 136.
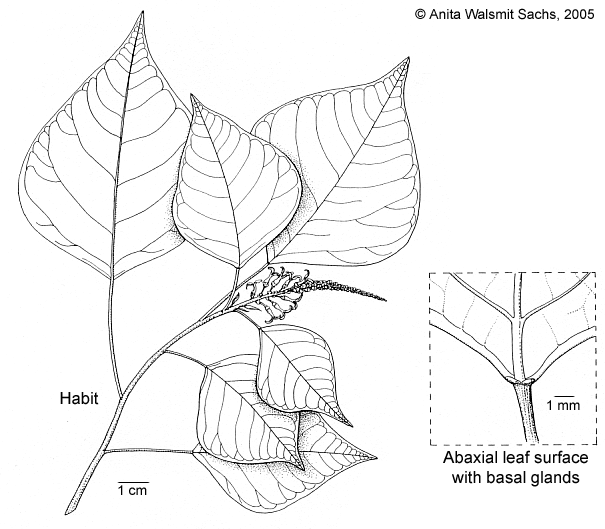
Tree up to 10 m tall, dbh up to 18 cm, with slender, terete, crooked to straight trunk, numerous spreading, flexible branches, and a flattish but spreading crown; branching „with KORIBA's model"; without buttresses. Glabrous. Bark pale brown to grey, non-fissured but lenticelled and mottled with pale patches, with greyish lenticels, yellow on the inside, very thin, soft; cambium yellow; inner bark pale whitish to purple. Wood tasteless and odorless or with a slight disagreeable odour. Stipules 0.8–2 cm long. Leaves: petiole 1–15 cm long, glandless; lamina orbiculate to ovate to lanceolate, 3–22 by 1.5–20 cm, index 0.8–4.6, base rounded to slightly emarginate to, rarely, cuneate, not or indistinctly (up to 1 mm) peltate, very base often attenuate, apex acuminate, lower surface usually whitish with larger veins of different colour, rarely not whitish at all, side veins in 9–15 pairs below the apex, angle of divergence 50–60(–80)°, partially to completely joined towards the margin, tertiary veins percurrent, quarternary veins reticulate and usually indistinct, adaxially without any prominent gland, but often with 2 glandular auricles up to 0.6 mm long, abaxially with 1–3 glands on each half of blade, 0.2–0.5 mm in diam. and close to the margin (5–15 mm distant from margin), basal ones sometimes enlarged (0.75–1.5 mm in diam.) and close to or touching the midvein if not absent. Inflorescences 3–30 cm long, usually bisexual, occasionally wholly staminate, staminate part 6–9 mm in diam. Bracts of staminate cymules c. 0.75–1.5 mm long, with a pair of large undivided glands 0.5–1.5 mm long and only slightly (0.2–0.3 mm) overtopped by the bract. Staminate flowers (1–)3 per cymule; pedicel 0.6–3 mm long; sepals 2, c. 0.6 mm long; stamens (6–)8–10 per flower with filaments c. 0.4 mm long and anthers c. 0.3 mm long. Pistillate flowers 0–4(–21) per thryse; pedicel 3–12 mm long; sepals 2, soon caducous; ovary c. 2 mm long, bicarpellate, papillate, style c. 0.6–1.5 mm long, stigma 1.5–3(–6) by 0.6–0.7 mm, apically undivided to slightly emarginate to shortly divided (see note 3), glandular over its whole length or only the apical or basal 0.5 mm glandless. Fruits 2–4(–8) per infructescence; bract persistent; pedicel 1–2.5 cm long; calyx caducous; fruit excl. style 4–9 by 5–8 mm, papillate, slightly sulcate, not carinate, style 0.3–1 mm long, stigma 1.5–3 by 0.6 mm; regularly opened fruits not uncommon, opening primarily loculicidally, pericarp c. 0.2–0.3 mm thick (i.e. c. 1:15–1:20 of fruit length), remaining columella c. 4 by 1 mm, slightly alate. Seeds c. 3.5–6 by 3–4 mm, upper half envelopped by arillode.
Distribution — Endemic to Malesia: Malay Peninsula (from the S border of Thailand/Isthmus of Kra southwards, incl. Singapore), Sumatra, Java, Borneo, Philippines (excl. N Luzon), Celebes, Lesser Sunda Islands (W part), Moluccas.
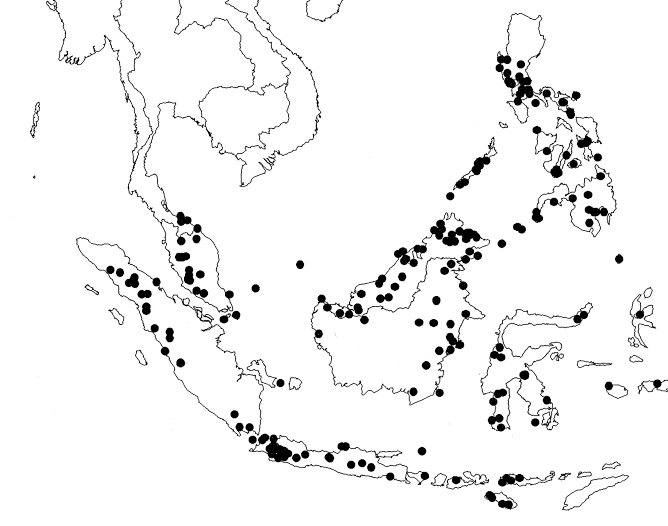
Habitat & Ecology — Found in secondary vegetation with Dendrocalamus, very open to old and shady secondary forest, light thicket, in Casuarina equisetifolia forest, disturbed Lithocarpus forest, at edges of lowland rainforest and Agathis forest, on roadsides, clearings, steep slopes (even extremely steep ones), forested ridges, exposed edge of hill forest, forest and secondary vegetation along river and in open rocky stream bed, on exposed cliff top, among heathlike vegetation, in waste land among high weeds, in cattle pasture, in 5-year-old Gmelina plantation. Soil: over granitic or sandstone rock, limestone rubble, shallow soil on rock with sulphur, ultramafic soil, silt clay, yellowish brown sandy loam, white sand, deep humus layer over clay, peaty soil, rusty red soil with concretions of iron, fertile compact soil. Locally scattered to frequent and gregarious, often one of the characteristic plants in recently cleared areas. Altitude: sea level up to 3,000 m (see note 4). Flowering and fruiting all year round. Evergreen (Smith, 1910). Ants' nests are commonly observed in the upper branches.
Uses — The fruits are used medicinally for wounds, e.g., in Sabah. The leaves are dried on the fire and are used after birth by putting them on the stomach, or the heated leaves are applied by the Malays to the abdomen for fever (Burkill, 1935). The bark and the leaves are, in connection with a certain kind of mud, used for dying clothes blackish. The dried and powdered leaves act as a vermifuge when fed to cattle (Heyne, 1950). The pounded leaves mixed with ashes and pepper sprayed in rivers act as fish poison. The wood is of low value and perishes quickly, but is good firewood. Sometimes planted, but suitable for afforestation only on better soils (Lörzing 223).
Vernacular names — Malay Peninsula: ludai paya, moyia (Ridley, 1924); mahang di-makan pelandok, mumah lapan (Burkill, 1935); maya maya, memaya, mouse deer's poplar (Corner, 1940). Sumatra: apasapas (N Sumatra); balanti; bodi (Minangkaban); baleh-baleh angin (Padang); delenge (Enggano); doelpok, sindoelpok, kajoe sindoelpok (E coast); karembi (Djasilin); kilimijah (Djambi); meloe (Gunung Koerintji). Java: balik lar (Gunung Muria); djarak prati, karembi, karembi badak, mroewoe, toetoep, toetoep lakek (Smith, 1910); kareumbi (Sunda); karumbi burrum (Gedeh); toetop(a)bang (Kediri area). Borneo: bangki (Bajau); belok malok (Dusun Keningau); bubalong, bungalau (Sungei); buta buta (Malay); butah butah paya, sedaman (Brunei); buta lalat (Martapura); delamato (Dusun Kandasan); gela (E Kalimantan, near long lees); kalang matoh (Dusun Tambunan); kepelak (Kenyah); ketapang lalat (Ulu Anap, Tatau, Sarawak); laru (E Kalamantan, near Longbagun); ludai api (Melaya); majin (Boengoeran Isl.); merendang (Bidayuh); tapang lalat (Iban). Philippines: balalanti (Cebuano); balanti, malabalanti (Salvosa, 1963; widespread); banti (Tawi-Tawi); batangig (Sibuyan); binuang-susong, malabinunga (Bataan; Merrill, 1912); bota-bota (Tayabas), botang gubat; buntatoli (Palawan); luglugun malagti; moguing (Bukidnon); santi (Yakan). Celebes: balante, tangu. Lesser Sunda Islands: lente, lente boé (Flores); woelloeroe (Sumba).
Notes — 1. The original type citation had been somewhat misleading. Geiseler (1807) wrote 'Habitat in Zeylona. Dahl'. Indeed, the original specimen in C does not give any locality and bears the note `Dahl a Dr. Thunberg'. A probable isotype is found in the Thunberg herbarium in Uppsala under no. 22680, as judged from microfiche 1036-951-24. It is filed under Croton sp., what explains the treatment in Geiseler’s Croton monograph. H. populneus obviously has never been found in Sri Lanka, and Geiseler must have been wrong in this respect. Thunberg collected in Malesia, but included also other collections from severel contemporaries into his herbarium. So, the real collector of the type remains uncertain.
2. Carumbium populifolium Reinw. was published first (1823) with a Latin footnote: `Carumbium: arbor tricocca, Crotonibus affinis'. Van Steenis (1959) considered this not as scientific description and Carumbium populifolium Reinw. as a nomen semi-nudum `which should not serve for purpose of priority'.
3. The concepts of the several Philippine varieties are somewhat overlapping and confusing. The Philippine plants of H. populneus are indeed remarkable by their often apically emarginate to divided stigmata. Pax & Hoffmann (1912) founded their var. siccus primarily on this character, but based its name on Excoecaria sicca Blanco, which is an Alchornea (Merrill, 1912). The stigmata, however, are not always divided on the Philippines and, very rarely, may be divided also in W Malesia. Smith (1910) cites this feature for Java, and three collections with distinctly emarginate stigmata were found on Borneo (e.g., Murata et al. B-560). Therefore, no taxonomic weight is given to this character.
4. Plants from Celebes often differ in the leaves which are narrowly elliptic to lanceolate (index 1.1–4.6 on Celebes, outside of Celebes 0.8–1.3) with short petioles, not whitish beneath, and sometimes very small flowers and fruits. H. sulawesianus is based on these differences. Usually, the differing plants were collected in elevations of 1,700–3,000 m, whereas outside of Celebes the species is only known from lower than 1,700 m. But no morphological gap to typical plants exists on Celebes, and even on high altitudes typical, broadleaved specimens were collected (e.g., Eyma 603, index 1.4). Therefore, H. sulawesianus is not kept separate from H. populneus. Although a comparable variability in leaf characters is not known from any other species of Homalanthus, it is not infrequent in other genera of the same tribe (Esser, 1994).
5. H. populneus is not known from Ambon. Frutex excoecans Rumphius, which was attributed to it by Merrill, Interpr. Herb. Amboin. (1917) 327, is probably identical with H. novoguineënsis.
Homalanthus remotus Esser, Blumea 42 (1997) 457, Map 10. — Type: Milliken 1178 (L holo; iso K), New Guinea, Irian Jaya, near Ilamik (1 hr walk E of Pronggoli).
Homalanthus leschenaultianus auct. non (Geiseler) Pax: Ridl., Trans. Linn. Soc. London 9 (1916) 147.
Tree up to 5 m tall. Glabrous. Stipules 0.5–1.5 cm long. Leaves: petiole 1–4 cm long, glandless; lamina narrowly ovate, 4.5–11.5 by 3–7 cm, index 1.6–2.7, base obtuse to rounded, very base often attenuate, not peltate, apex subacuminte to acuminate, lower surface whitish with darker larger veins, side veins in 10–14 pairs below the apex, angle of divergence 55–65°, distinctly to indistinctly joined towards the margin, tertiary veins reticulate to percurrent, quarternary veins reticulate, adaxially with a basal pair of pillow-shaped glands 1.2–1.5 mm in diam., abaxially with a basal pair of conspicuous, 0.8–3 mm long glands 0–4 mm distant from the midvein, on each side of midvein with additional 3–5 smaller, 0.3–0.4 mm long, submarginal glands 2–3 mm distant from the margin. Inflorescences 1.5–5 cm long, bisexual, staminate part 3–5 mm in diam. Bracts of staminate cymules with a peduncle 0.5–0.8 mm long, with a pair of undivided glands 0.3–0.5 mm long and only slightly to not overtopped by the bract. Staminate flowers 3 per cymule; pedicel up to 0.5 mm long; sepals 2, 0.3–0.6 mm long; stamens (4–)6 per flower, with very short filaments and anthers 0.3–0.4 mm long. Pistillate flowers 1 per thyrse; pedicel 8–11 mm long; sepals 2, c. 0.5 mm long; ovary 1–1.5 mm long, bicarpellate, papillate, style 0.5–1.5 mm long, stigma 1.5–3 mm long, undivided, with an apical to subapical gland c. 0.7–1.5 mm long. Fruits 1 per infructescence; pedicel 17–20 mm long; calyx persistent; fruit excl. style 4–5 by 4–5 mm, slightly sulcate, slightly carinate at back, papillate, style persistent; opened fruit not seen. Seeds not studied.
Distribution — New Guinea (Irian Jaya).
![]()
Habitat & Ecology — Found in Nothofagus dominated forest, and on limestone ridges. Altitude: 1,500–1,900 m. Flowering and fruiting: February, March, May, June, August, October.
Use — The wood is used for making fire by friction.
Vernacular names — New Guinea: song, songkal (Jali).
Note — This new species is unique in the genus by the pedunculate floral bracts. The name refers to this character. It is obviously closely related to H. arfakiensis and differs from the latter also in the adaxially always glandular leaves without an acute base, reticulate tertiary veins and larger abaxial glands, and by larger stigma glands.
Homalanthus trivalvis Airy Shaw, Kew Bull. 21 (1968) 415; Esser, Blumea 42 (1997) 458, Map 4. — Type: BSIP (Whitmore) 2236 (K holo; iso L), Solomon Islands, Santa Ysabel, Maringe Lagoon, Buala.
Homalanthus xerocarpus Airy Shaw, Kew Bull. 33 (1979) 536. — Type: Kajewski 1691 (BRI holo; iso A), Bougainville Isl., Kupei Gold Field.
Tree up to 17 m tall, girth up to 60 cm, crown large, diffuse; without buttresses. Glabrous. Bark light brown to grey, pustular; inside chestnut brown. Slashwood with a smell `like medicine'. Stipules 1.2–4 cm long. Leaves: petiole 3–13 cm long, apically with an abaxial-lateral pair of disc-shaped glands 0.3–0.75 mm in diam.; lamina orbiculate to ovate, (3–)4.5–13.5 by (3–)4–14 cm, index 0.9–1.5, base rounded to emarginate, very base often slightly attenuate, usually not peltate, but rarely peltate by up to 6 mm, apex acuminate, lower surface whitish with complete venation of different colour, side veins 8–12 pairs below the apex, angle of divergence 45–50°, at least partially joined towards the margin, tertiary veins percurrent, quarternary veins reticulate to indistinctly percurrent, adaxially glandless, abaxially with 0 or 1 laminar gland c. 0.25 mm in diam. and c. 5 mm distant from leaf margin, very rarely also with the otherwise petiolar glands partially or totally on the lamina. Inflorescences 4–10 cm long, bisexual but pistillate flowers often absent, staminate part c. 6 mm in diam. Bracts of staminate cymules c. 1 mm lang, with a pair of undivided glands 0.5–0.75 mm long and distinctly overtopped by the bract. Staminate flowers 1 per cymule; pedicel 1–1.5 mm long; sepals 2, c. 0.5 mm long; stamens c. (6–)9–12 per flower, with filaments and anthers each c. 0.25 mm long. Pistillate flowers 1 or 2 per thyrse if not absent; pedicel 2–6 mm long; sepals 3, c. 0.5 mm long; ovary 1.5 mm long, tricarpellate, papillate, style 1–2 mm long, stigma 2.5–4 by 0.25 mm, undivided, with an apical gland 1–2 mm long. Fruits 1 or 2 per infructescence; bract often persistent; pedicel 2–4 cm long; calyx persistent; fruit excl. style 8–15 by 8–13 mm, tricarpellate, papillate, distinctly to indistinctly sulcate, each carpel carinate, style persistent; opening regularly and loculicidally, hardly along the septa, pericarp c. 1–1.2 mm thick (i.e. c. 1:10 of fruit length), each septum with 3 vascular strands, remaining columella c. 1 mm wide, not alate. Seeds 4.5–5 by 3.5–4 mm, totally covered by an arillode.
Distribution — Bougainville and the Solomon Islands.
(circles on Bougainville and Solomons)
Habitat & Ecology — Found in well-drained secondary forest, cloud forest, on steep slopes, ridge tops, cliffs, or flat plains. Soil: coral ground, lime. Altitude: sea level up to 1,700 m. Locally common. Flowering and fruiting all year round with a maximum in October.
Uses — Pounded leaves are used for stupifying fish. Wood charcoal is used as paint on canoes. A drink made from the plant shall rid people from evil spirits.
Vernacular names — Bougainville: coondou, navi, tim-bar-ci. Solomon Islands: hi'isikima, nonilu, nonumba, sikima, sikona (Kwara'ae).
Notes — 1. The citation of this species for New Guinea by Airy Shaw (1980a) is erroneous and was based on a specimen of H. longistylus with some tricarpellate flowers (see note 5 under H. longistylus).
2. H. xerocarpus is united here with H. trivalvis. Its type is a small-leaved form with quite indistinct leaf glands and only 6 stamens per flower, collected in a cloud forest at higher elevations, but reveals no substantial difference with H. trivalvis. Airy Shaw himself, when describing H. xerocarpus, did not cite any significant difference with H. trivalvis, but compared the species only with the Polynesian H. acuminatus which is quite different indeed.
Homalanthus polyadenius Pax & K.Hoffm. in Engl., Pflanzenr. IV, 147, xiv, add. VI (1919) 57; Airy Shaw, Kew Bull. Add. Ser. 8 (1980) 119; Esser, Blumea 42 (1997) 459. — Type: Ledermann 11948 (holo? B, †), Papua New Guinea, Schraderberg, Gebirgswald.
No extant type of this species could be traced. The description of Pax & Hoffmann (1919) gives only few distinguishing characters. Noteworthy are an undivided adaxial-petiolar leaf gland, a pair of large abaxial laminar leaf glands near the leaf base, a row of additional submarginal leaf glands, and a pair of oblong glands at the floral bracts; staminate cymules with 3–5 flowers, and 5–6 stamens per flower. These characters are not sufficient to allow a determination of the specimen. It is improbable that H. polyadenius is a distinct species. It may be synonymous either with Homalanthus nervosus or with H. populifolius. The comments of Airy Shaw (1980a), who stated that H. polyadenius may be a juvenile form of H. vernicosus (= H. nervosus) may be right but are hypothetical. In my opinion, no definite synonymy should be proposed, and H. polyadenius should not be interpreted.